生命:复杂性的新阶段
“生命是什么?”1943年,物理学家埃尔温·薛定谔(Erwin Schrödinger)在都柏林的系列讲座中提出了这个问题。薛定谔的回答具有极大的预见性,因为他在我们对生命有正确的遗传学基础认知之前,就给出了答案。他指出:我们能够像解释物理学、化学那样科学地解释生命。但同时他也明白,我们不能仅仅靠查阅一份清单来为生命下定义。像所有的复杂事物一样,活的生物体控制着大量的能源与物质流,所以它们一定具有某种新陈代谢的形式。它们吸收并排泄着能量与营养。而且与从龙卷风到水晶的复杂然而无生命的诸多实体一样,它们也会再生。因此,新陈代谢或者再生这两个概念,都无法单独对生命做出一个令人满意的定义;只有二者在一起共同发生作用,从而创造一种新层次的复杂性才是问题的关键。因此,薛定谔建议另寻方法来定义生命的独特性。生命不仅是复杂的——它比宇宙中的其他任何事物都更为复杂;由于宇宙总体趋向于无序,因而生物所能够达到的有序性就非常令人注目了。“在生物体的生命周期中所显示的事件都表现出令人赞叹的规律和秩序,这是我们所能见到的任何无生命的物体所根本无法比拟的。”[1]
恒星可以在热力学的下行电梯上向上攀升(参见附录二),但是生物体向上攀升的手法更为敏捷。实际上,埃里克·蔡森认为,通过计算维持有生命的生物体抗拒热力学第二定律破坏压力的能量流密度,我们可以大致然而客观地测量出生物体所达到的复杂性的等级。[2]表4.1列出了蔡森所说这些能量流的近似值。表的右列,是在一定时间内经过一定质量的自由能量的流量值,其结果表明生物体可以不间断地处理比恒星更为密集的能量流。正是这种能力,使它们能够在热力学下行电梯上攀升得更远更快。薛定谔有一句名言,每一个生物体似乎都拥有令人吃惊的“不断从它周围环境中吸收秩序”的能力。[3]最简单的结构也许存在的时间最为长久,越复杂的结构出现的时间越晚,这表明后者的形成是一项更加困难的进化工作。最后,位于图表下半部的那些复杂事物则显然更为脆弱。恒星和行星可以存在数十亿年,最长寿的生物(至少就我们所知)也只能活几千年,大多数只能存活几天或几年。最复杂的结构消亡如此之快,这也是生物处理超密集能量流的困难程度的一种衡量尺度:这正是生物顽强抵抗热力学第二定律所付出的代价。因此,讨论生命也就是讨论复杂和秩序的新层次,即以自身更加脆弱为代价,获取控制和组织自由能量的新能力。正如马丁·里斯所说:“恒星比昆虫简单。”[4]但恒星存在的时间更为长久。
表4.1 自由能量速度密度估算表
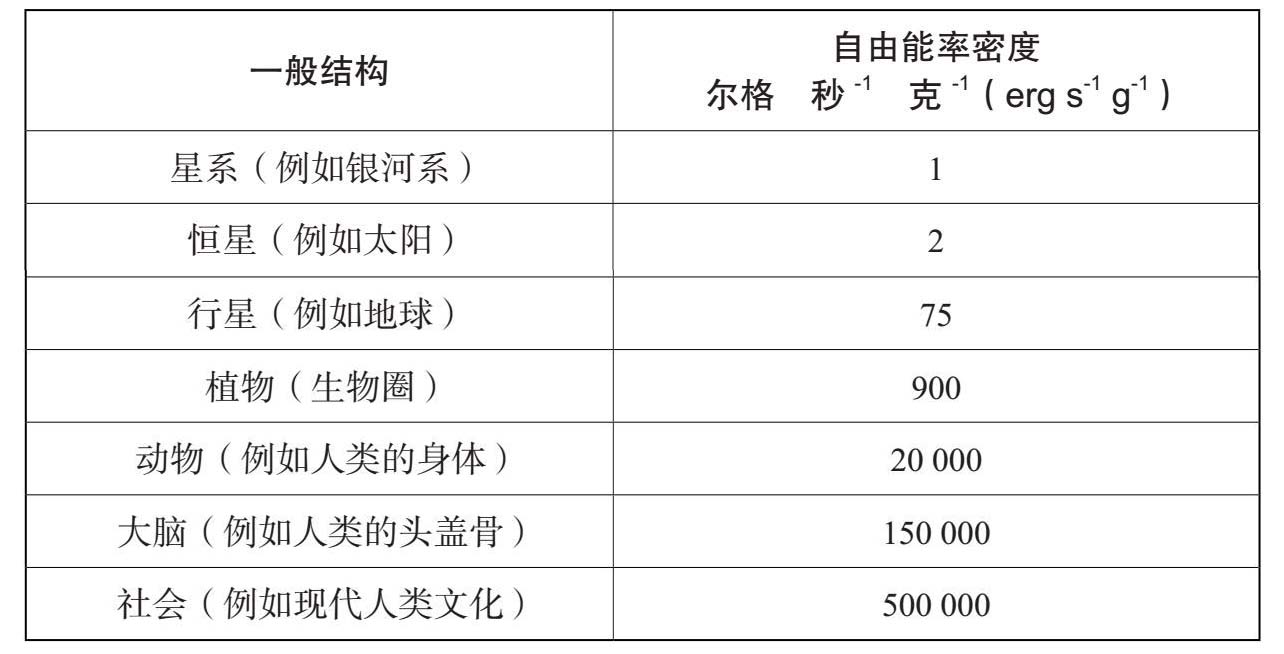
资料来源:埃里克·蔡森:《宇宙的演化:自然界复杂性的增长》(马萨诸塞,坎布里奇:哈佛大学出版社,2001年),第139页
化学过程或许在宇宙的其他地方也创造了生命,尽管目前我们并不知道这是否属实。但在地球6亿年的诞生过程中,生命的确出现了。从地质学的标准看,由于早期地球是一片不毛之地,生命的出现可谓神速。从生命出现的那一刻起,活的生物体就开始繁殖,新的生命形式开始在无尽的序列中不断变化,这种变化令人眼花缭乱,每一种生物体都细致地调整自己,以处理周边环境中特有的能量和资源。与恒星或水晶这些普通的、无目的的负熵构造不同,在与熵展开更为灵活的游击战争的过程中,生物体能够不断适应新的地形以及新的挑战。生物以集体为单位探索环境,这种方法不见于无生命世界。它们所发现的是新的能量源以及新的自我组织的方法,以便在能量的暴风骤雨中存活下来。不是所有的变迁都导致更大的复杂性,但有些变迁确实能够做到。这就是生命为什么有如此惊人的能力,可以魔术般变出各种新的复杂性的原因。
维持这些复杂实体的不同能源从何而来?答案很明确:最根本的源头就是引力。我们知道,正是引力创造了恒星——密度和温度极高的物体。但宇宙总体上讲是很冷的,其平均温度为绝对零度以上3℃,即宇宙背景辐射的温度。所以恒星如同数十亿个散发着光和热的小点一样镶嵌在这寒冷的宇宙中。我们拥挤地住在一颗恒星的附近,我们能感觉到从太阳中心核熔炉中释放出的巨大能量流正源源不断地注入太空,这绝非偶然。
至于活的生物体的复杂性的规则,它们与在天文学尺度中起主导作用的规则有所不同。个体生物(至少我们已经知道的)是在比行星或恒星小得多的尺度内繁荣起来的。在这些较小的范围内,引力是重要的,但其他一些力更为重要。生命形式主要是由控制原子运动的电磁力(electromagnetism)与核作用力(the nuclear forces)决定的。这些力决定了原子如何聚集又如何构成更大、更复杂的分子。
但是在复杂性的生物学层面上,新的规则也出现了。活的生物体按照独特的、开放性的变化规则运作,这些规则叠加在较简单、较具决定论意义的物理和化学规则之上。生物学规则之所以会出现,在于活的生物体的繁殖过程具有非常高的精确性。处理巨大的能量流是一项细致的工作,它需要极为精确的结构,创造和再创造这种结构的规则必然是复杂而精确的。繁殖系统如果只是近似地复制生物体的机制,那么很快就会丧失其必需的精确度(当然,如果繁殖系统的复制过程精确到完美的程度,那就排除了任何变化的可能性)。因此,高度精确的新陈代谢需要一个高度精确(但并不完美)的繁殖。这就是为什么像我们人类这样的大型活的生物体需要比细菌更多的遗传学信息。同样,这也是为什么大多数关于生命起源的研究更侧重于遗传密码源头的原因,遗传密码这一复杂的分子级“软件”,解释了为什么活的生物体能够以其他复杂实体所无法企及的精确度进行复制。
总之,从化学到生命的转变是宇宙史上的一个巨大转变。按照精确的新蓝图所复制的复杂生物体引入了一种历史变迁的新类型——当然这发生在地球上,或许也发生在宇宙中的其他地方。化学物质的组合形成了活的生物体,而要解释新出现的生物属性,我们却无法简单地通过分析构成生物体的化学成分来实现。因此,要了解生物,我们就需要一个新的范式,一个能带领我们超越核物理学、化学或地质学规则,而进入生物学领域的范式。本章将讨论生物发展变化的基本规则以及关于生命起源的一些新观点。重点将放在查尔斯·达尔文(Charles Darwin)的观点上,他第一个明确阐述了生物学变化规则的独特性。下一章将研究地球生命的历史。
达尔文和进化论
造物主或神灵以某种方式把生命带入这个原本没有生命的世界,许多社会用以解释生命的就是这类假设。现代科学认为,这种解释是一种方便法门,因为仰仗神灵的理论或多或少可以解释任何事物而不需要客观的证据。相反,现代科学试图把生命的诞生解释为各种无生命的力与过程所造成的结果,正如它对太阳和地球的形成所做出的解释一样。
现代生物学用以解释生命起源和发展的基本观点就是通过“自然选择”(natural selection)而展开的“进化”(evolution)。该理论由查尔斯·达尔文于1859年在他的著作《物种起源》中第一次做了系统阐述。[5]达尔文很少使用进化这一术语,或许因为它似乎暗示着某种驱使生物朝特定方向变化的神秘力量,因而会与他自己的观点相互矛盾,他认为生物的变化是一个开放性的过程。然而,极力推广这一术语的赫伯特·斯宾塞(Herbert Spencer)看到,生物变迁是由“低级”生命形态(life-form)向“高级”生命形态的运动,是一种不断进步的形式。这样的观点是不合时宜的,因为这种研究会将武断的、主观的价值判断引入我们对生物体发展历史的理解。尽管有这么多由进化所引发的联想,我们仍将使用这个术语,因为它最适用于达尔文关于生物变迁的理论。
达尔文论证道,物种并不是一成不变的实体。它们处在不断的变迁之中,而且变迁的方式受到一些简单规则的控制。一个物种就是一大批个体生物,它们彼此有足够的相似性而能够杂交,但相互之间又不完全相同。物种是根据这些个体生物所共有的特征,而不是它们之间微小的差异而定义的。但经过很长一段时间,个体特征的随机变异引发整个物种总体特征的改变。比如,其平均高度也许会改变,或者其大脑的平均容量会增加。经过数千代的积累,这些微小的变迁最终必将改变整个物种的总体特征。我们想要了解物种如何变迁,就必须了解一些个体特征为什么以及如何变得越来越普遍,与此同时,其他个体特征则在衰退乃至消亡。
达尔文知道,在大多数种群之中只有少数个体才能存活至成年、繁衍后代。然而正是那些能够存活和繁殖的个体决定了该物种的未来。所以代代相传的生命仅仅是那些幸存者的后代。(进化就像历史一样,显然是由胜利者书写的。)但是,哪一个能繁殖,哪一个不能繁殖,这又是由什么所决定的呢?当然,也许一切只是偶然而已。但是达尔文认为,从长远来看,最有可能得以存活和繁殖的是这样一些个体,它们幸运地继承了其父辈能够稍稍更好地适应周围环境的特征。接着,它们会把这些相同的特征遗传给后代。岁月流逝,这些特征就变得越来越普遍,因为不具备这些特征的个体只能繁殖较少的健康后代,直到它们这一脉全部消亡。历经数千代,许多诸如此类的微小变化将确保物种不断变化或进化,以便更好地适应周围环境。打个比方,我们可以说是环境“自然地”选择了某些特征,而舍弃了其他一些特征,就像饲养动物的人“有意”选择这样一些而不是另外一些动物去喂养。正是在这样一种比喻的意义上,随着时间的流逝,物种就渐渐“适应”了它们的自然环境。
适应,是现代生物学中一个极其重要的概念,我们应当更加仔细地对其下一个定义。它是指一切生物看上去都与其所生存的环境精确适应的事实。的确,生物与周围的环境是如此完美的适应,以至于达尔文的反对者辩称,诸如人类的眼睛或大象的鼻子等器官必定是由一位慈祥的造物主所设计的,而至今仍有人这么认为。达尔文试图证明,无目的的自然过程同样也能够做到这点。适应有助于解释生物的高度多样性,因为生物所要适应的环境是复杂多变的。要描述这些不同的环境,生物学家和生态学家使用了栖息地(habitat)和生态龛(niche)这两个概念。栖息地只是物种生存的地理环境。生态龛这一说法要复杂一些,它还包括了物种生存的方式。niche一词起源于拉丁语,意为“巢穴”。在建筑学中,niche指墙上放置雕塑或其他物品的壁龛。在生物学和生态学中,生态龛是指生物在进化过程中被塑造或逐渐适应的特定的生活方式。啄木鸟的生态龛是由在树上发现可食用的昆虫的方式而确定的。在包括我们人类在内的大型动物的内脏中,许多单细胞细菌找到了颇具吸引力的生态龛。但是,环境当然也会改变——当环境改变之际,旧的生态龛就会湮灭,而新的生态龛在其他地方形成。由于环境各不相同且不断变化,生物想要生存就必须不断适应环境。这就是进化为什么永不停息的原因。完美或“进步”并没有固定的标准,因此适应是一个永无止境的过程。
现代生物学家用进化的观点来解释地球上生命形式的多样性。他们尝试用同样的观点来解释地球生命的起源,因为无生命的物质似乎也可以通过简单形式的自然选择而进化。如果有适宜的环境和足够的时间,它们最终会进化为活的生物体。什么是生命?它又是如何改变的?进化的观点是理解这些问题的基础,因此在开始叙述地球生命的历史之际,我们必须更详尽地描述这一理论,也必须了解它是如何从各种关于生命起源的陈旧解释中演变过来的。
现代进化论的起源
我们知道,在17、18世纪,一些欧洲的科学家开始怀疑犹太教——基督教《圣经》中的创世神话。《圣经》好像说物种是上帝在大约6000年前创造的,并且基本保持着上帝造物之际的本来面貌。这种信仰甚至得到18世纪现代分类学(taxonomy)体系或生物学分类的创始人、瑞典植物学家卡尔·林奈(Carl Linnaeus)的支持。然而,即使在17、18世纪,这种观点也难以自圆其说。比如,许多化石显示从未在《圣经》或历史记载中提到的奇怪生物的存在。一些海洋生物化石出现在了形成于数百万年前的高山上,或者被深深地埋在了岩石中。当然,化石所在的地点说明了它们一定是在数百万年前就埋葬在这里的。
所有农民都知道这么一个事实:狗、猫、牛以及羊等物种并不是一成不变的。事实上,通过仔细的选择交配,鸽子或者狗的育种家能够培育出一些非常奇特的动物。达尔文也是伦敦两家鸽子俱乐部的会员,对于育鸽人的活动,他非常着迷。达尔文描述了他所看到的一些鸽子,很显然,它们都是从普通的野鸽驯化而来的:
从英国传书鸽(English carrier)和短面翻飞鸽(short faced tumbler)的比较中,可以看出它们在喙部之间的奇特差异,以及由此所引起的头骨的差异。传书鸽,特别是雄鸽,头部周围的皮具有奇特发育的肉突;与此相伴随的还有很长的眼睑、很大的外鼻孔以及阔大的口。短面翻飞鸽的喙部外形差不多和鸣禽相像,普通翻飞鸽有一种奇特的遗传习性,它们密集成群地在高空飞翔并且翻筋斗。侏儒鸽(runt)身体巨大,喙粗长,足亦大;有些侏儒鸽的亚种,项颈很长;有些翅和尾很长,有些尾特别短。……突胸鸽(pouter)的身体、翅、腿特别的长,嗉囊异常发达,当它得意地膨胀时,令人惊异,甚至发笑。[6]
这些奇特的动物与上帝最初的创造属于同一物种吗?或者它们完全就是新物种?如果它们是新的,那么上帝显然在不断地修补物种——这种修补似乎暗示上帝最初的创造并不完美。自哥伦布之后,欧洲人绵延数个世纪的远航探险,也使他们意识到有很多物种在《圣经》中并没有提到。太平洋、美洲和欧亚地区的动植物,彼此之间存在着许多差异,这对基督教神学家而言是一个巨大的挑战。是上帝创造了所有这些物种吗?如果是,那物种为何会如此纷繁芜杂?上帝又为何如此奇怪地让它们在全世界随意分布?为什么英国没有袋鼠,而澳大利亚没有熊猫?
到18世纪后期,一些生物学家一直在思考,随着时间的流逝,生物是否可能按照某种自然机制而改变,因为似乎很难设想上帝会频繁地修改他的作品。也许这样一种机制可以解释,为什么会有那么多的物种和亚种,为什么有那么多的物种在《圣经》的创世叙述中没有记载。问题是谁也解释不了物种为什么变化,又如何变化。
19世纪早期,达尔文的叔叔伊拉斯谟·达尔文(Erasmus Darwin)提出,为了更好地适应环境,物种是会进化的。这说明了问题,因为一切现有的生物看起来的确非常适应它们的环境。但是,与同时代所有的生物学家一样,他也不知道物种是以什么方式来适应环境的。在1809年出版的一本书中,法国的博物学家让——巴蒂斯特·拉马克(JeanBaptiste Lamarck)提出了一条可能的机制。动物通过自己的努力而得到的一些微小变化也许会遗传给它们的后代。他举了这样一个例子,长颈鹿的祖先伸着脖子去吃高处的树叶。其中伸得最努力的那几只就把长脖子遗传给了后代。慢慢地,长脖子就变得越来越普遍了,最终成了整个物种所独有的主要特征。然而很不幸,任何一位动物育种家都能指出这一理论的谬误。后天习得的特征——即通过独特的生活方式或特殊个体的努力而获得的品质——一般是不会传到下一代的。只有遗传特征才以这种方式传递。我们在健身房花费的时间并不能保证我们的孩子会健康。一头肥猪并不一定会生出肥硕的小猪,但是如果这只猪的祖辈肥硕,那它很有可能会生出肥硕的后代。
拉马克提出的机制并不说明问题。但如果生物的遗传构造由它们的过去(即它们从父辈那里所继承的)所决定,那它们怎么能适应现在的生存条件?这就是达尔文所要解决的难题。达尔文从小就被动物所吸引,20多岁的时候,他已经成为一名专业的博物学家。像那时大多数的博物学家一样,达尔文明白物种很容易受外界的影响。他也明白人类可以通过人为的选择而改变物种。但是他不明白在没有人类干预的情况下,为何物种也会发生变化。除了上帝和人类,还有什么能使一些个体生存繁衍,而使另一些个体走向消亡?
1831年,以博物学家的才华和某种幸运的家庭关系,达尔文获得“比格尔”号(Beagle)帆船上的博物学家一职,这艘船正在进行环球探险。旅途中所见到的丰富的物种,以及关系密切的物种之间细微的变化,都使达尔文惊讶不已。在例如犰狳等生物的身上,他目睹了进化的清晰的化石证据。在南非,他看到了与活的动物十分相似却又有着细微差别的动物化石。然而正是在远离智利太平洋海岸的加拉帕戈斯岛上,他找到了最终使其形成进化论的线索。在那儿,在一些新近形成的火山岛上,他发现了一些似乎与美洲大陆有着密切关系的雀鸟。然而各个岛上的雀鸟并不完全相同。例如,它们的喙部就有一些细微的差异,这些差异使每一种物种与它们所居住的岛屿上那些兴盛的特殊植物与动物极为适应。
这就是伊拉斯谟·达尔文所说的那种适应性的明确证据。很显然,物种在某种意义上可以“适应”环境的变化,但它们是如何做到的?大约在1838年,达尔文读到了现代人口学先驱托马斯·马尔萨斯(Thomas Malthus)的著作,似乎启发了他理论的核心观念。马尔萨斯注意到,在包括人类在内的大多数物种中,很多个体(有时候是大多数)都无法繁衍后代。在达尔文看来,显然只有那些能够繁衍的个体才会对后代的性质产生影响。因此,为什么一些个体能幸存下来,而另一些却不能?搞清楚其中的原因很重要。当他研究了一些育鸽人的行为之后,答案就很明确了,育鸽人会有意识地挑选一些个体让它们繁衍后代。在达尔文的时代,这已经发展成为极其成熟的手艺:
关于鸽子,最有经验的育种家约翰·西布赖特爵士(Sir John Selbright)曾经说:“他可以在三年的时间内培育出特定的羽毛,但要花六年的时间才能得到他想要的头和鸟喙。”在萨克森(Saxony),人们充分认识到自然选择的原则对于美利奴羊(merino sheep)的重要性,以至把选择当作一种行业:把绵羊放在桌子上,研究它,就像鉴赏家鉴定绘画那样;在几个月期间内,一共举行三次,每次在绵羊身上都做出记号并进行分类,以便最后选择出最优良的,作为繁育之用。[7]
但是自然界中的选择是怎样的?是什么“选择”一些个体使之繁衍,而使另外一些走向消亡?能够繁衍的个体与不能繁衍的个体之间有什么差别?
达尔文认为答案就是“适应性”(fitness)。在统计学意义上,那些幸存下来并繁衍出健康后代的个体比未能存活和繁衍的个体要略微健康一些。它们之所以能够繁衍后代,就是因为它们健康,可以生存得更为长久,并且有健康的配偶。当然,在一些个别的案例中,运气可能会起作用(如果遭受电击而死亡,那么无论你再怎么“适应”也不重要了)。但是对于大多数个体而言,在很长一段时间内,适应必定起了举足轻重的作用。就平均水平而言,那些活到成年并繁衍后代的个体肯定要比未能存活的个体稍许健康、稍许适应环境一些。因此,并不是物种真正地适应;恰恰相反。那些纯粹由于偶然机遇能更好地适应环境的个体最有可能存活下来,并为该物种的后代定了型。
达尔文认为,这种以随机统计学过程展开的挑选,会在一切形式的生命中世代发生,如果重复的频度足够,它就能像人类的饲养者一样有效地改变物种。这样的过程一再发生,每一代都会发生数百万次,环境淘汰了一些个体而让其他一些得以幸存。后代只遗传这些幸存者的特性,结果,久而久之整个物种变得越来越像这些幸存者。打个比喻,环境扮演了动物育种家的角色。这就是为什么达尔文称这种机制为“自然选择”,而称动物育种为“人工选择”(artificial selection)的原因。
通过这种方式,达尔文指出,这种不断重复的纯粹统计学的、完全无意识的过程,可以解释物种是如何在不知不觉中不断地适应环境变化的。要完全理解达尔文的观点,关键在于理解其中大多数过程具有随机性特点。个体与其父母的差异是细微的,但是这种差异本质上是随机的。不管怎样,它们不是“试图”去适应。“进化”的不是个体,而是物种的总体特征。
达尔文论证道,经过长时间的反复,这些机制也能解释不同物种是如何起源的,因为,很显然,某个物种的种群分布很广,并且为环境的细微差异所影响,可能以稍微不同的方式进化。在达尔文看来,加拉帕戈斯岛的雀鸟就是不同物种形成的早期阶段的典型例子。达尔文论证道,在经过相当长时间之后,这样的过程可以解释在地球上所有生物的多样性。作为地质学家查尔斯·赖尔的仰慕者,达尔文确信地球存在了很长时间——也许,如此漫长的时间足以使得微小的变化最终形成了现有物种的极大多样性。
这是令人震惊的结论,因为它们意味着一场彻底的革命:地球上所有美丽和复杂的生物,从变形虫到大象,再到蜂鸟,乃至人类本身,都是由这种盲目的、重复的过程所创造的。看来无意识的过程不但能够创造行星和银河系,甚至还能创造生命本身。[8]这样的推理似乎剥夺了上帝存在的一切理由,这也是达尔文的理论直到如今仍遭到强烈反对的原因。
以下是达尔文本人对自然选择的工作方式所做的描述:
没有明显的理由可以解释,为什么在驯化状态下如此有效的原理竟然不能在自然状态下发生作用……(既然)诞生的个体比可能存活的更多。天平上的毫厘之差便可决定哪些个体将生存,哪些个体将死亡——哪些变种或物种将增加数量,哪些将减少数量或最后灭绝……由于这种生存斗争,不管怎样轻微的,也不管由于什么原因所发生的变异,只要与其他生物或者外部自然,处在无限复杂的关系里面的个体或者任何物种有利,就会使这样物种保存下来,并且一般会遗传给后代。后代也因而有了较好的生存机会,因为任何物种的许多个体,周期性诞生,但是其中只有少数能够存活。我把每一个有用的微小变异得以保存下来的这一原理称为“自然选择”。[9]
通过自然选择进化的证据
在《物种起源》一书中,达尔文尽可能清楚阐述他的进化论观点。他还试图应付对进化论可能提出的一些异议。对于他所负使命的艰难程度,他几乎不抱什么幻想。他的大多数读者都是传统的基督徒。他们相信上帝创造不同的物种,物种可能由盲目过程而产生的想法令他们感到震惊。因此,达尔文大多数的观点是针对这些读者的。
根据化石记录,他也能够证明物种是随着时间变化而变化的。但是这也可能是一个最没有说服力的论据,因为这些化石记录包含着一系列的个体,即已变成了化石的“照片”;反对者可以轻松论证,说每一个个体都是上帝分别创造的一个不同物种,而现在已经灭绝了。达尔文不得不证明,存在着变迁的物种。一些化石看起来的确是现存动物类型的中间物种。其中最有名的化石就是生活在1.5亿年前的始祖鸟,一种像鸟一样的恐龙。始祖鸟的样子看起来既像是爬行动物,又像是鸟类。第一个化石样本于1862年被发现,恰好在《物种起源》出版之后,达尔文还可以在以后的版本中对此做出评论。这正是达尔文的理论所预言的那类发现。但对于反对者而言,也能轻易地指出那些化石记录是极不完善的。在整个化石谱系中还存在着巨大的鸿沟,因此,要想证明自然选择的原理,化石记录本身永远也不能够提供完全令人满意的证据。
达尔文还提供了证明其理论的其他一些类型的证据。他认为,现代物种所拥有的很多共同特征,标志着它们是从共同的祖先进化而来的。很奇怪,这一主张最有说服力的地方就在于,一些明显毫无用处的特征通过某种生物学上的守旧性而从遥远的祖先那里保存了下来。鲸鱼有指骨,在今天它们已经毫无明显的适应性功能。但是指骨的存在让我们设想,现代鲸鱼是从陆地动物演变而来的,它们的手和手指曾经有过实际的用途。事实上,现代鲸鱼可能是早期也同样适应水栖生活的哺乳动物河马的远亲。自然选择的理论能够很容易地解释这种现象,因为它意味着生物是以很缓慢的步伐进化的,还保留着许多过去的特征,即使这些特征已经没有任何实际用处。这一论据是很有说服力的,因为传统理论难以解释这类证据。为什么上帝在设计每个物种的时候却保留了这些无用的器官?
达尔文还能够证明,物种的地理分布比较符合他的理论而不是神创说。为什么上帝不把某个物种安置在地球上所有它们能够适应的地区呢?为什么不是所有沙漠都有骆驼满地跑呢?为什么博物学家反而发现,在一个特定区域内,大多数物种之间都有密切的关系呢?举例来说,澳大利亚的鼠类生物与袋鼠之间的关系就比欧洲的老鼠更为密切。当然,达尔文的答案是有袋类动物生活在澳大利亚,因为那里就是它们祖先生活的地方,那里也是它们进化的地方。
批评达尔文的人尤其厌恶其理论的另一层含义:人类可能是猿的近亲(这层含义在今天依然活跃如初;参见第6章)。对于某些人来说,此种想法至今仍使人讨厌;人类学家伊夫·科庞(Yves Coppens)曾记得他的祖母对他说:“也许你是一只猴子变的,但我绝对不是。”[10]但是在达尔文的时代,他的理论还面临许多其他困难。例如,大多数地质学家确信,地球的存在还不足1亿年。达尔文知道物种的自然选择需要相当漫长的时间,才能在地球上产生像现在这样种类繁多的生物,他承认仅1亿年的时间是不够的。他相信进化的进程极其缓慢。事实上,他深信这一过程如此之缓慢,以至于根本无法直接观察得到,因此所有进化论的证据都是间接的。此外,19世纪的生物学家并不真正明白遗传是如何发生作用的。达尔文的理论要发生作用,那么遗传的机制必须十分精确(否则不会有稳定的物种存在),但也不能太精确(否则将永远不会发生变化)。重要的是,父辈的品质要能够传递给后代,但是同样重要的是,一些细微的差异也许会增强,也许会威胁某个个体的健康。但是,由于谁也没有充分了解遗传机制是如何运行的,因此谁也不能确定遗传是否严格按照所需的方式来运行。达尔文只能说,父辈的品质是“混合的”,就像两种颜色混在了一起。但是这似乎必然导致在生物的每一代中,生物范式所发生的变异——哪怕是有益的——都会被抹杀掉,其结果是连自然选择本身也变得不可能了。对遗传学缺乏确切的认识削弱了达尔文理论的可信度,这一过程长达半个多世纪。
达尔文所面对的大多数难题在20世纪才得到解决。宗教对进化论的抵制减少了。同时,支持这一理论的新证据也出现了,而且该理论的空白点也得到填补。达尔文理论在一些重要方面得到修正和改进。结果,达尔文的中心论点成了解释地球生命历史的现代科学基本原理。
达尔文理论如今之所以被广泛接受,是因为在20世纪可以直接观察到进化的作用。研究像果蝇这一类繁殖很快的小物种,是最容易观察到进化的过程的。新形式细菌出现以回应抗生素的使用,我们也可以从中看到进化的作用(在下文将作进一步的探讨)。
化石记录也比达尔文时代要丰富得多,而且新的发现对于进化的长期性做出了更加完整的解释。虽然这些解释不能完全证实达尔文的观点,却与之十分相符。现代的年代测定技术把地球的年龄从1亿年向前推到了40亿年,为达尔文的进化过程提供了一个比原先要长40倍的时间。20世纪的生物学家最终逐步了解了遗传是如何发生作用的,而且他们的解释完全符合达尔文的理论。与达尔文同时代的格里格·孟德尔(Gregor Mendel)早就指出了遗传的基本原理,尽管他的著作在20世纪之前一直没有受到重视。他指出,尽管有性繁殖的生物从双亲那里遗传了特性(或者说是基因),但这种遗传处在一种分散的状态——一些特性从父系这里遗传,另一些特性则从母系这里遗传。他还指出,在很多实例中,这些特性往往只有一个会在后代身上得到体现。如果你父母的眼睛分别是蓝色的和褐色的,这并不意味着你的眼睛会是蓝褐色的,你只会遗传一种颜色。所以遗传并不会导致令达尔文感到害怕的特性的衰微。特殊的基因也许不会遗传给所有的后代,但如果遗传了的话,那就是一种完整的传递。我们还确切地了解了基因是怎样一代代遗传下去的。脱氧核糖核酸即DNA,将遗传信息从生物体极精确地传递给它们的后代,因此物种具有极大的稳定性。但这一过程并不是完美无缺的。当脱氧核糖核酸在复制自己的时候,平均在每十亿个遗传信息中可能会出现一个错误,其概率相当于一名打字员在输入50万页文字中出现一处错误。在进化过程中,如此微小的变异是难以避免的。[11]因此,弗朗西斯·克里克(Francis Crick)和詹姆斯·沃森(James Watson)于1953年对脱氧核糖核酸结构的解释,对于巩固达尔文理论,使之成为现代生物学的中心观点是至关重要的。
现代微生物学还证实了达尔文的另一个基于直觉的想法:地球上所有的生物体都是有联系的。所有活的生物体,从最简单的细菌到最大型的现代哺乳动物,都包含使用同样基本化学过程和途径的细胞,而且它们都使用相同的遗传密码。所以说,所有活的生物体都是有联系的。这意味着以下二者当中只有一个成立:要么生命仅仅进化了一次;要么生命不止进化一次,但是这些尝试中,只有一个世系得以存在到今天,而其他世系最终都消亡了。不论哪个成立,从人类到香蕉再到海鞘和变形虫,当今所有的生物体都来自同一个(细菌)祖先。
达尔文的理论在一些细微之处已经略有修正。例如,达尔文似乎相信所有由进化而引发的变异之所以发生,是因为它们增加了个体生存的机会。但现在证明许多遗传变异的发生是随机的。有大量的遗传物质(或许占人类基因组的97%)对成年个体的结构是没有影响的。所以这方面的变异不会直接影响个体的生存机会。其普遍原理似乎是这样的:对于物种个体的生存机会没有影响的随机变化,可以导致整个物种的遗传结构(或基因型)发生缓慢的、本质的变化。但是,如果有的时候一些不活跃基因被重新激活,则这些“中性的”变异在未来也许会变得特别重要。
达尔文似乎相信,进化的速度是稳定的,但现在证明这显然是不正确的,在气候或环境稳定的状态下,物种可能变化得很慢。但是当环境或气候迅速变化,物种会进化得很快且趋向于多样性。现代细菌恰恰就是这样来应对抗生素的挑战而不断进化的。当抗生素被广泛使用的时候,受抗生素影响最小的细菌个体有很大可能性会突然开始繁殖更多健康的后代。在几代之内,它们的基因将会支配整个物种。以这种方式出现的细菌新种或多或少都对抗生素有了一定的免疫力。如今这样的节奏在进化的过程中看来是正常的。在地球的历史上,既有进化变异非常迅猛的时期,也有生物相对稳定的时期。按照“间断进化”(punctuated evolution)的理论,进化时快时缓。这一理论由尼尔斯·埃尔德里奇(Niles Eldridge)和斯蒂芬·杰·古尔德(Stephen Jay Gould)于1972年提出。
地球生命的起源
现代达尔文理论可以解释地球早期的简单生命形式是如何进化成现代生物的(见下一章)。但能进一步解释最初生命形式的起源吗?对于生命起源于无生命的物质,能否做出纯粹的科学解释呢?至少在古希腊时期,科学家真诚地接受生命可能发乎自然的观点。[12]理由是相当充分的。毕竟,动物尸体上的蛆虫似乎就是出自乌有之乡。在17世纪,通过使用最新发明的显微镜,研究证明空气中充满微小生物,假设它们的卵在空气中传播并在腐肉上寄居,这样就可以解释蛆虫一类生物所谓的自发产生。但这并不能排除微生物自发产生的可能性,也许生物可以通过飘浮在空中的某种形式的“生命力”而产生。
在一次极其简单的试验中,法国生物学家路易·巴斯德(Louis Pasteur)似乎终于证明了生命自然产生的观点不能成立。所有的生物都是以有机分子为基础的——也就是说,这些分子的主要成分是碳。因为碳元素能以复杂的方式结合在一起,它可以形成比其他任何元素都要复杂且有着较大差异的分子。到19世纪,许多实验表明,饱含有机物的汤,在煮沸杀死所有生物体后,若放置在密封容器中就不会出现任何生命。但也有人认为这是因为容器隔绝了所有的生命力。1862年,巴斯德做了一项很有独创性的试验来证明这个观点。他煮了一锅饱含有机物的肉汤,然后把它放在并不封闭的鹅颈瓶里,与空气接触。一方面,如果生命力真的存在,它一定会进入容器并在有机物的基础上产生新的生物;另一方面,空气中飘浮的孢子(spore)或微生物将不能到达瓶子的颈部。巴斯德的肉汤中没有出现生命,这个实验延续了一个多世纪,今天在巴黎仍然可以看到那瓶肉汤。巴斯德的实验似乎最终证明了生命并不能自然产生,而且空气中也没有飘浮着的生命力。生命只能产生自生命本身。
这破解了一个谜团,不料又出现了另一个谜团。假如生命不能由无生命的物体产生,那么早期地球上的生物又是如何出现的?古生物学家知道,生命大约在寒武纪(Cambrian Era)经历了一次大爆发,我们现在知道这发生在距今不到6亿年。生命突然迸发,这又如何解释呢?难道生物学家要回到神创论吗?许多19世纪的生物学家确实回到了神创论,因为任何关于生命起源的纯科学解释都假设生命产生于无生命的物质,而巴斯德似乎已经证明这是不可能的。另一种可能的解释认为,有机分子与无机分子之间存在极大的差异。也许它们的区别就在于它们是否能产生生命的能力上。是碳元素有什么特殊之处吗?19世纪中期,化学家证实,在实验室中无机分子可以合成为有机分子,显然这一理论也是不成立的。
直到20世纪,一个更为合理的关于生命起源的科学理论出现了。这一理论由亚历山大·奥巴林(Alexander Oparin)和约翰·伯登·霍尔丹(John Burdon Haldane)于20世纪20年代首先提出,即用进化论的基本观点来解释生命的进化以及地球生命的起源。其主要观点是,就某种程度而言,甚至在复杂然而无生命的化学物质中,进化就已经在发生作用了。因此,甚至某些化学物质也可以进化,比如水晶,它们自身保持着稳态且会生长出新的水晶体。当水晶生长之际,那些繁殖出最稳定的“后代”(也就是说这些后代最能适应周围的环境)的化学物质,要比后代不能很好存活的化学物质生长得更为迅速。这个过程与达尔文的进化论极为相似。当这些化学物质变得更加适应周围环境的时候,它们也会变得更加复杂,直到我们最终把它们看作是活的生物体。生物学家将这一过程称为化学进化。
化学进化是如何产生最早的活的生物体的?答案依旧是不明确的。要想理解这些困难,我们必须把问题分成几个层面。首先,我们需要解释生命的基本原材料如何生成:这是化学的层面;其次,我们需要进一步解释这些简单的有机物怎样聚合成更为复杂的结构;最后,我们还需要解释在所有生物中都存在的脱氧核糖核酸(DNA)精确繁殖机制编码的起源。目前,对于第一个问题我们已经有了较为满意的答案;对于第二个问题我们的答案还似是而非;而我们对于第三个问题仍然百思不得其解。
第一项任务现在看来真是出奇的简单,生物大多由碳和氢构成,由于碳元素极具活性,所以是最关键的。再加上氢、氮、氧、磷以及硫,这些成分总共占了生物净重的99%。[13]事实证明,只要条件适宜,这些化学物质充足,就很容易制造出简单的有机分子,其中包括氨基酸(合成蛋白质的材料,所有生物的基本结构材料)与核苷(合成遗传密码的材料)。[14]巴斯德的实验似乎表明了这样的有机分子不可能自然形成。现在我们知道了其中的原因:今天的大气中含有大量的自由氧,对于有机分子而言这是一个极为恶劣的环境。氧是很活泼的元素,当它起反应的时候会产生热量(我们通常都会意识到它在火灾中的破坏力)。对于像木头、纸张这一类物质的有机分子,氧具有很大的破坏性。一有可能,在缓慢燃烧的形态下氧气会把它们分解成水和二氧化碳。
但是,奥巴林和霍尔丹指出,在早期地球的大气中几乎不存在自由氧。也许生命是在寒武纪之前,在一个没有氧气的大气中出现的,如此就确保了简单的有机分子长期生存,使得化学进化所必需的复杂而缓慢运动的化学过程得以发生。1952年,这种可能性被两位美国科学家哈罗德·尤里(Harold Urey)和他的研究生斯坦利·米勒(Stanley Miller)所做的一项十分著名却又特别简单的实验所证明。他们在一个巨大的密封曲颈瓶中注入甲烷、水和氨,制造了这样一个早期大气的模型。然后,加热混合剂,用通电的方式输入一些自由能量,来模拟早期地球上空必然存在的闪电。7天之后,他们在曲颈瓶中发现了暗红色的软泥。其中包含20种最重要的氨基酸中的若干种。氨基酸是简单的有机分子(大约由20—40种原子构成),它们以不同的模式链接在一起,形成在有机物及所有生物结构中占主导地位的蛋白质。实验在略有不同的条件和略有不同的仿真大气中重复了多次,科学家发现所有20种基本氨基酸都能用这种方式制造出来。这样,我们就拥有了制造蛋白质这一生命基本构造材料的基础。但是“米勒——尤里实验”也创造了少量的其他一些重要的有机分子,其中包括糖和构建遗传密码分子的核苷的主要成分。
有人宣称米勒和尤里已经接近创造活的生物体了。但现在证明事实并非如此。在创造简单的有机分子与创造生命之间,还有许多艰难的步骤。无论如何,早期地球大气中存在的氨和甲烷可能要比两位化学家所设想的要少,而二氧化碳和氮比他们所设想的要多。在这样的大气中,简单的有机分子并不会很多。但不管怎样,“米勒——尤里实验”还是相当重要的,表明早期地球上组成生命的许多基本化学构造的产生并不是那么艰难。
自这个实验以后,氨基酸、简单的核苷,甚至构成细胞膜的磷脂(phospholipid),在许多令人惊异的地球和太空的环境中都有所发现。在星际空间的尘云中已确定有氨基酸存在。还发现了大量的水和酒精,这些对于磷脂的产生也是至关重要的。我们知道,在陨石与彗星中都有水蒸气和许多简单的有机分子。太空中存在水以及各种简单有机分子,这意味着在整个太阳系的历史上,或是经由猛烈的碰撞,或是通过行星表面不断飘移的宇宙尘埃,太阳系一直受到生命原材料的轰击。事实上,现在看来在太阳系中的若干星体,包括火星、金星、木星的卫星——木卫二和木卫四,以及土星的卫星——土卫六上曾经有过(有些至今仍有)液态水,因此它们本来应该进化出简单的有机分子,即使现在它们都是一片不毛之地(金星和火星看来都是如此)。还有可能的是,至少在这些行星诞生之初,由于陨石的撞击而切割下来的岩屑在它们中间飘浮,曾经彼此交换过有机物。例如在1996年,有人声称在南极洲发现一块陨石,它大约在1.3万年前坠落于地球,其中包含着的气体与火星稀薄的大气中的混合气体相同。假如这一说法是准确的(许多科学家对此持怀疑态度),这就意味着这块陨石很可能来自火星。如果在火星上发现了生物,那么它们与我们之间应该是有关系的,对于这种可能性我们不得不加以重视。[15]总而言之,现在看来形成生命的基本化学物质在早期太阳系中似乎是很丰富的。
第二项任务要困难一些。它将解释这些最多只包含着几十个分子的简单化学物质,是如何组成生命存在所必需的巨大而复杂的结构的,甚至连病毒都包含大约100亿个以特别形式组成的原子,而每一个植物和动物的合成细胞都包含1万亿——100万亿(即1012——1014)个原子。这种在规模和复杂性方面的巨大飞跃是在什么地方、以什么样的方式达到的呢?目前还无法确定。而正是这种变化把有机化学物转变成了真实的生命。生命最早起源于何处?这个问题的答案目前有三种可能。最初或可能产生于太空,或在行星表面,或在行星内部(这是最近提出的一种可能性)。
许多年以来,弗雷德·霍伊尔(Fred Hoyle)和钱德拉·维克拉马辛格(Chandra Wickramasinghe)一直在论证,是外来生命在地球上洒下了种子。这种理论被称为胚种论(Panspermia)。如果生物体最早是在宇宙中其他地方的行星上产生的,那我们仅仅是把这个问题转换到了另外的行星上,我们仍然要解释在那里生命是如何被构造出来的。此外,也有人认为极其简单的生物是在太空合成的。我们知道,在太空确实会有化学过程发生,而且一些简单有机物强壮到足以能够经过漫长的太空旅行而存活下来。但是现在看来,似乎生命本身不大可能起源于太空,那里的能量和原料供应不足,因此使化学过程一般发生得非常缓慢。除此之外,对生命至关重要的许多化学反应都离不开液态水,而在太空中并未发现液态水的存在。
行星——那里的环境更为复杂,自由能更为充裕,液态水可以采集,所产生的化学物质密度更大,品种也更为丰富——为生源论(Biogenesis)提供了一个更有希望的发展空间。直到最近,大多数生物学家还在假设,如果生命诞生于地球,那它一定出现在地球的表面。早在1871年,达尔文就曾说过,生命可能肇始于“某些温暖的小池塘,里面含有各种氨和磷盐,再加上光、热、电等因素”。[16]在早期地球的海洋中,或海边的某个地方,自然的化学和物理过程是如何把化学物质合成为简单生物的?从达尔文时代起,生物学家就在尝试解答这个疑问。
在所有这些理论中,水扮演了至关重要的角色。氨基酸和核苷一旦形成,只要它们在水里,在某种程度上就能得到保护。氨基酸和核苷的分子可以自然地排列成长链,尽管这个过程需要更干燥一些的环境。这样的链条也许就是在海岸边的浅水塘中形成的,在这些池塘里,分解的分子周期性地干涸,然后再分解。在适宜的条件下,氨基酸链形成蛋白质,而核苷的链形成核酸。于是在数百万年之后,早期地球的海洋中就充满了简单的有机物,它们能够组合成更复杂的模式。A. G.凯恩斯——史密斯(A. G. Cairns-Smith)曾指出,在浅水中,黏土的微小晶体也许为更复杂分子的形成提供了一个模板。[17]在这里,静电力可能吸附由黏土分子结构控制的复合原子,直到它们开始以新的方式联结在一起,甚至早期细胞中的黏土晶体还可能通过提供可反复使用的模板,以产生用于其主细胞新陈代谢所需的化学物质,这无疑扮演了现代脱氧核糖核酸的部分角色。
不管早期的有机分子是如何形成的,它们能够形成一种由简单的蛋白质、核酸以及其他有机分子组成的有机的薄“汤”。这些分子有一种倾向,形成由磷脂构成的简单薄膜或“皮肤”,并凝结成微小的液滴,“就好比辣椒酱里面的油滴”。[18]一些分子可以通过它们的皮肤吸收化学物质,其过程与进食有些相似,这为它们提供了自身扩张以及进一步吸收化学物质的能量。而且,当这些分子长得过于庞大笨拙后,就分裂成好几个部分,而每一部分都会以自己的方式独立发展,就像油脂表面上的一颗大水滴会分裂成小水滴一样。因此,40亿年前的海岸以及温暖海水里面,也许早就存在着具备很多生命活动特征的有机分子。它们组成有着表层皮肤的细胞状液滴,它们“吞食”其他化学物质,它们分成不同的液滴,就像繁殖一样。
所有这些理论好像都有几分道理,但都不能解释生命从无到有的整个过程。关于“温暖的小池塘”说还存在着一些问题,早期大气可能不像米勒和尤里所假设的那样适宜于有机物的进化——尤其是如果正如某些证据所表明的那样,最早的生物出现于38亿年前,而那时地球表面正频繁地遭受着来自宇宙空间物体的撞击,那么情况就更加如此了。最近的研究为解决这一问题提供了新的途径,它揭示了先前不为人所知的细菌形态——原始细菌(archaebacteria)的存在,它们是在地表之下生长进化的。[19]就像所有的原核生物(prokaryote,最简单的单细胞生物)一样,原始细菌没有细胞核。但原始细菌不像大多数原核生物那样从阳光或其他细胞中汲取能量,而是以地球上的化学能“为生”。它们可以从铁、硫、氢以及许多其他埋藏在岩石中或溶于海水里的各种化学物质中汲取能量。原始细菌可以舒适地生活在地表以下很深的地方,甚至可以承受极大的高温与高压,因此它们的存在表明生命可能起源于行星地下而不是表面。20世纪90年代,人们发现原始细菌生活在地表以下1000米的岩石中;人们还发现它们不但生活在温度超过沸点的海床火山岩口,而且还生活在海床以下的渗水岩中。2001年,在被研究者形容为海床上“消失的城市”的一大片区域里,也发现有原始细菌。在那里,热量并不是由火山活动产生的,而是来自一种名为橄榄石(olivine)的绿色岩石与海水之间发生的化学反应。这样的区域在地球早期历史上是很普遍的。[20]但是原始细菌在地球表面也大量存在。研究发现在怀俄明州黄石国家公园的温泉里就有原始细菌。最后,它们在极端的地方生活,说明在邻近的行星和月亮上也可能存在或曾存在过生命,因为在太阳系的其他地方也有类似的栖息环境。
原始细菌比大多数现代生物更适合作为地球生命的最初形式,这一理论模型能够得到多个理由的支持。原始细菌生活在自冥古宙以来鲜有变化的环境中。在地球早期历史上,周期性的陨石撞击极为普遍,足以毁灭临近地表的一切生命,而原始细菌能够在地表以下生活,意味着它们基本没有受到陨石撞击的影响。在臭氧层形成之前,地表还保护了原始细菌免受地球大气变化和轰击原始地球的强烈紫外线辐射的威胁。尽管喜热的原始细菌的栖息地在我们看来也许是可怕的,但是它可能为早期生物的形成提供了一处最佳场所。这些环境包含有大量的化学养料,可以制造出“米勒——尤里实验”中形成的各种有机物。尤其是在炽热的火山口周围,这些环境包含有可以引发若干化学反应的大量能量。对原始细菌的遗传基因材料的研究也表明,它们的进化比起大多数其他幸存的有机物要慢得多。也许最令人惊奇的是,所有最古老的生物,不论是原始细菌还是普通细菌都是耐热的。这表明无论我们怎样为地球上最早的有机物分类,它们也许只是在深海火山口周围极其肥沃的环境下得到进化的喜热的有机物而已。如果这些论证是正确的,那么在地表或靠近地表的寒冷环境中能够生存的新物种诞生之前,生命很可能就最早已经出现在地表之下和大海洋之中了。[21]
第三项任务是要解释遗传密码的起源,这比前两项任务更难处理。从某种意义而言,这是所有问题中最基本的,因为所有现代生命形式的关键似乎就是核苷的分工问题,核苷储存并阅读制造有机物(基因组)的指令,而蛋白质则运用这些指令构成一个生物体。大体而言,核苷操纵复制的过程,而蛋白质掌控新陈代谢。它们之间的差别就好比电脑的硬件与软件。因此,到底哪一个进化得早一些呢?是新陈代谢(化学活动)还是复制(遗传密码)——或者是它们同时进化?[22]
奥巴林的理论暗示新陈代谢出现得较早,而复制机制进化得较晚。这种观点一眼看上去也是似是而非的,就好比没有软件硬件可以存在而不是相反。在奥巴林的理论中,最早的生物是一些能以粗略的方法繁殖甚至“进化”的化学物质,而且它们进化的方式很复杂。许多人很难理解这种随机的过程是如何产生复杂性的。进化过程,即便是粗略的进化过程,事实上也不完全是随机的。在早期地球上的随机化学作用中,有些会形成更为稳定的副产品。因此早期分子的聚合过程并不是每次都从头开始的。相反,相对稳定的分子每次产生,都有可能幸存下来,从而依次成为下一步试验的基础。正如切萨雷·埃米利亚尼所指出的那样,对于一只猴子而言,即便它用了数百万年的时间随意敲击打字机键盘,要打出一部《圣经》的概率几乎为零。但如果加上这么一条规则,即每敲击出一个正确的字母,就将它固定在那个位置上,那么概率就完全不同了,我们可以期待在10年之内完成一部《圣经》。[23]或者用另外一种方式表达,我们可以说,早期地球的有机化学物就已经服从于进化的法则了。大多数化学物质消失了,而适应了环境的那些化学物质很可能被“固定下来”。极少数存活得足够久远的,就得以生息繁衍,此后的世代就都是它们的族裔。环境就以这种残酷方式“选择”了那些最有能力生存繁衍的化学物质。
这种化学进化也许是一种极其强大的变化机制,许多理由支持这一假设。例如,一些科学家推测,存在一种深层机制,它促进我们意料之中的具有纯粹的随机性特点的组织的形成。[24]在某些类型的化学反应里,一个特定的化学物质也许会催化或激励它自身的形成,这一过程被称为自身催化(autocatalysis)。当这种类型的足够量的化学物质聚合在一起,最终可能会发生失控反应(runaway reaction),就好比把临界质量的铀填装在一枚炸弹里面一样。一旦达到临界质量,化学链的反作用会很快形成极为复杂的结构。如果这一逻辑(从本质而言是数学逻辑)成立,则意味着作为最早生命形式的大型的、复杂的化学物质的构成也许是有机化学的自然趋向。如果是这样,那么凡是在宇宙的任何地方,只要条件允许大量有机化学物形成并且相互作用,生命的出现就几乎是可以确定的事情了。
但是所有形式的进化都需要其复制机制达到足够的精确度,否则,即便是最成功的特性也会随着时间的流逝而消失。因此,即便是把新陈代谢置于首位的理论也必须解释最初存在着怎样的复制机制,这可并不容易。当今(除一些病毒以外)一切生物遗传密码的关键就是脱氧核糖核酸,它是一种极其复杂的分子,包含数十亿个原子。如果解开人类的一个脱氧核糖核酸分子,其长度将近2米。脱氧核糖核酸的原子以很精确的模式排列在一起,就像一份软件那样,包含创造一个生物所需的全部信息。我们身体里的每一个细胞都有这样一套完整的指令,尽管它只使用了脱氧核糖核酸指令手册中极小的一部分。指令手册根据特定的环境选择并激发相应的指令。因而,脑细胞和骨细胞可以分别使用指令的不同部分。
每个脱氧核糖核酸分子都有两股核苷的长链,中间有横档相连,就像梯子一样。这架梯子又扭曲成一条长螺旋,好似螺旋楼梯。每格梯子都有四种名为碱基的简单原子中的一种附着其上。每种碱基只能与其他四种碱基中的一种联结,因此梯子的每根横档都是由两种碱基以严格的序列联结。腺嘌呤(简称A)只能与胸腺嘧啶(简称T)联结,而胞核嘧啶(cytosine)(简称C)只能与鸟嘌呤(guanine)(简称G)联结。这些出现在梯子双股上的碱基序列中包含制造构成生物体之蛋白质的密码。每三个一组的碱基为一个氨基酸编码。特殊分子定期解开脱氧核糖核酸螺旋的一段,读出由三个核酸组成的序列。这些氨基酸在细胞的其他地方组成链条,以便制造出大量促发化学反应并形成细胞结构的蛋白质。脱氧核糖核酸也可以自我复制。首先,当构成梯子的每一根横档的两个碱基彼此分离,整个双螺旋线就像拉链一样从中间一分为二。然后,每个碱基从周围环境中吸引一个与自身类似的配对物,A与T、C与G相结合,直至最初那个螺旋的每一半都构成一条全新的互为补充的链条。每个脱氧核糖核酸的分子以这种方式可以形成两个新的分子,它们或多或少都与原先的分子相一致。
这一复杂、精细而雅致的机制是怎样产生的呢?解释这个问题是现代生物学理论所面临的最具挑战性的任务之一。一个难题是脱氧核糖核酸自身似乎是无能为力的。就像所有的软件一样,没有硬件,脱氧核糖核酸无所作为。因此很难想象它是如何独立进化的。而新陈代谢(即“硬件”)最先进化的观点也有问题——尤其让人难以理解的是,简单进化过程如何能产生高级的复杂事物。如果细胞复制并不那么精确的话,埃米利亚尼所描述的机制将不能正常运作。即使是复杂生物体的进化,其计划蓝图也可能在以后的世代渐渐变得模糊。正因为如此,许多致力于解决这些问题的人坚持认为,如果没有更为精确的复制能力,生命就不能获得新的重大的复杂性。这又导致我们回到了这样一个观点上来,尽管这个观点同样困难重重,却认为遗传密码先于复杂的新陈代谢。
20世纪60年代,脱氧核糖核酸的近亲核糖核酸(RNA)的发现推动了这些理论的发展,与脱氧核糖核酸相比,核糖核酸不那么无所作为。与脱氧核糖核酸一样,核糖核酸是软件,可以为信息编码。但核糖核酸只存在于一条链中,这意味着它也可以像蛋白质那样折叠起来,并参与新陈代谢活动。因此,在生命中核糖核酸扮演双重角色;它能自行繁殖,并提供一整套繁殖的指令。它既是硬件也是软件。最早实现足够精确的复制而具有一些“遗传”特点的分子也许就是核糖核酸制造的。事实上,甚至今天的一些病毒仍以核糖核酸而不是脱氧核糖核酸作为它们基因组的基础。
核糖核酸既可以做硬件又可以做软件那样活动,这一发现形成了一些主张核糖核酸乃是生命最早形式的理论。这些理论与曼弗雷德·艾根(Manfred Eigen)和莱斯莉·奥格尔(Leslie Orgel)的研究工作相结合,认为遗传密码最早进化,先于复杂的新陈代谢,甚至先于细胞。[25]不幸的是,核糖核酸不能像脱氧核糖核酸那样精确地复制自身,这就造成了真正的问题。一个好的但不至于非常好的复制系统也许是一切可能世界中最糟糕的,因为它可能坏到累积错误但又好到将错误忠实地遗传给后代。与生命起源的“新陈代谢优先”模式所需的比较草率的复制形式相比,这样一个系统也许会更快导致崩溃。(著名的核糖核酸拥护者曼弗雷德·艾根懊恼地把这描述为“错误的灾难”。)[26]
弗里曼·戴森提出,这两种理论或许可以综合在一起。[27]也许新陈代谢确实是最早出现的,而没有精确复制机制的细胞则支配着地球上的生命达数百万年之久,虽然有其局限性,仍设法推动那些至今在现代细胞中还在发生的各种新陈代谢过程的不断进化。其中一个至今仍然存在于所有生物之中的新陈代谢过程,便是在一个被称为ATP(三磷酸腺甙)的分子内储存能量。而这个过程的发生,与作为核糖核酸重要组成部分的另外一个分子有着密切关系。因此,核糖核酸也许就是在这样的细胞中进化的,它提供了一个比它们进化的外部世界更为适宜的环境。最后,就像某种寄生生物一样,核糖核酸劫持了细胞的繁殖机制,直至在一种早期的共生形式中,细胞与寄生于细胞之中的核糖核酸达成妥协,细胞致力于新陈代谢,而核糖核酸则致力于繁殖。由于有了比较精确的复制机制(如果“错误的灾难”多少能够避免),这些细胞中的核糖核酸最终就会进化成为核糖核酸的近亲脱氧核糖核酸。
也许正如我们所知道的这样,生命就是从这两种截然不同类型的生物体的共生状态中出现的,这两种生物体,一种擅长新陈代谢,一种擅长编译密码。这种分工至今仍然存在于细菌以及许多病毒中(参见第5章)。细菌常常利用类似病毒的自由飘浮的软件碎片为己所用,而病毒一类的实体则利用细菌和其他生物的新陈代谢能力来繁殖。我们可以想象,在早期世界,新陈代谢用类似病毒的器官来稳定其自身的复制机制,而病毒类的生物体则利用细菌完成它们的新陈代谢,直到二者最终结合为单一的生物体。
即使这些理论无一能够让人完全信服,我们也不必大惊小怪。生命起源的完整理论还不存在。对于解释遗传密码起源,亦即真正复杂生物体出现的关键问题,我们仍然处于相当困难的境地。但是,近几十年来的进步是迅速的,而正在进行的研究也将在下一个10年或20年间给大家一个更令人满意的解释。
本章小结
达尔文的进化论经过20世纪的修正,为现代生命科学提供了重要的基本理念。达尔文认为,物种哪怕一点点随机的变异都能解释,为什么一些个体要比其他个体更具有繁殖后代的可能性。那些能较好适应环境的个体更有可能存活至成年期并产下健康的后代,因而可以把基因遗传给下一代。这样,物种通过我们所称“自然选择”的方式而慢慢变化,长此以往,就形成了全新的物种。
在地球历史的早期,这样的过程也可能塑造飘浮在太空中、地球表面以及地表以下的有机化学物。这些有机物中的“成功者”幸运地存活下来,历经几十亿代之后,通过化学形式的自然选择,形成了更为稳定和复杂的有机化学物。在地球诞生后不到10亿年,最早的生物体就以这种方式被创造出来了。这些生命体是一切现代生命形式的祖先。
延伸阅读
达尔文是现代思想的为数不多的奠基人之一,其著作值得一读。《物种起源》(1859年;1968年再版)至今仍有较强的可读性,而斯蒂夫·琼斯(Steve Jones)在《新物种起源论》(2000年)中对达尔文的理论进行了一些更新和修正。阿曼德·德尔塞默的《我们宇宙的起源》(1998年)对生物和进化做了一个简明扼要的导论,还有许多其他现代教科书也是如此。近几年来,生物学界出现了一些针对非专业读者的优秀著作。斯蒂芬·杰·古尔德在进化方面的著作很值得一读,即使当他还没有取得主流地位的时候。丹尼尔·丹尼特(Daniel Dennett)的《达尔文的危险思想》(1995年)是一部现代经典作品,而厄斯特·迈尔(Ernst Mayr)的《冗长的论证》(1991年)和约翰·梅纳德·史密斯(John Maynard Smith)的《进化理论》(1975年第3版)观点略为陈旧。在介绍现代试图解释生命起源的优秀著作中,保罗·戴维斯的《第五奇迹》(1999年)是最新的和最通俗易懂的。埃尔温·薛定谔的《生命是什么?》(1944年,1992年再版)仍值得一读,他的观点被弗里曼·戴森的《生命的起源》(1999年第2版)一书所修正。A. G.凯恩斯——史密斯的《生命起源的七大线索》(1985年)和罗伯特·夏皮罗(Robert Shapiro)的《起源》(1986年)的讨论精彩而容易理解。关于强调细菌作用的生命观,林恩·马古利斯和多里昂·萨根则为我们提供了一个极具可读性的导论[“微观世界”;特别参见《微观世界》(1987年)。埃里克·蔡森[著有《宇宙的演化》(2001年)、《生命纪》(1987年)和《宇宙》(1988年)],斯图亚特·考夫曼(Stuart Kauffman)[著有《在宇宙的家庭中》(1995年)]和罗杰·卢因(Roger Lewin)[著有《复杂性》(1993年)]、《人类进化》(1999年第4版)]论述了关于复杂性的一些观点以及在现代关于生命的讨论中它们的作用。
[1] 埃尔温·薛定谔:《生命是什么?》,载《生命是什么?生命细胞的物理学特点》,以及《精神与物质》和《自传述略》(剑桥:剑桥大学出版社,1992年,1944年初版),第77页。
[2] 埃里克·蔡森称Φm为“自由能率密度”,而且他用“单位时间单位密度的能量单元”衡量自由能律密度;他补充说:对于这一概念,“就好像天文学家对于亮度——质量比,物理学家对于力密度,地质学家对于特定辐射通量,生物学家对于特定的新陈代谢比,工程师对于力——质量比(power-to-mass)那样熟悉”。《宇宙的演化:自然界复杂性的增长》(坎布里奇,马萨诸塞:哈佛大学出版社,2001年),第134页。
[3] 薛定谔:《生命是什么?》,第73页。
[4] 马丁·里斯:《探索我们的宇宙及其他》,载《美国科学》,1999年12月。
[5] 另一位博物学家阿尔弗雷德·拉塞尔·华莱士(Alfred Russel Wallace),差不多同时形成了类似的观点,而且他与达尔文一起于1858年首次在林奈学会(Linnaean Society)出版的定期刊物上发表了两篇论文,对这一理论进行了介绍。
[6] 查尔斯·达尔文:《通过自然选择的物种起源:在生存竞争中优势种类的保存》(哈蒙斯沃思:企鹅出版社,1968年),第82页,伯罗(J. W. Burrow)编辑并序。
[7] 达尔文:《物种起源》,第90页。译文引自周建人、叶笃庄、方宗熙译:《物种起源》,商务印书馆1995年版,第42页。——译者注
[8] 盲目的运算过程可以创造出独特的复杂性事物,丹尼尔·丹尼特在其新作中,将此描述为“达尔文的危险思想”。参见《达尔文的危险思想:进化与生命的意义》(伦敦:亚伦·莱恩出版社,1995年)。
[9] 达尔文:《物种起源》,第441—442、115页;引自蒂姆·梅加里(Tim Megarry):《史前社会:人类文明的起源》(巴辛斯托克:麦克米伦出版社,1995年)。译文引自周建人、叶笃庄、方宗熙译:《物种起源》,商务印书馆1995年版,第534、535、76页。——译者注
[10] 休伯特·里夫斯(Hubert Reeves)、约尔·德·罗斯奈(Joёl de Rosnay)、伊夫·科庞和多米尼克·西莫内(Dominique Simonnet):《起源:宇宙、地球和人类》(纽约:阿卡德出版社,1998年),第138页(科庞撰写的部分)。
[11] 阿曼德·德尔塞默:《我们宇宙的起源:从大爆炸到生命和智慧的出现》(剑桥:剑桥大学出版社,1998年),第135页。斯图亚特·考夫曼指出,有序与混乱之间那种微妙的平衡也许其本身就是进化过程的产物,这种可能性很吸引人。参见《在宇宙的家庭中:关于复杂性规律的研究》(伦敦:维京出版社,1995年),第90页。
[12] 这一部分材料基于戴维·布里斯科(David Briscoe)1989年以来在麦考瑞大学的演讲。参见林恩·马古利斯和多里昂·萨根:《生命是什么?》(伯克利:加利福尼亚大学出版社,1995年),第64—69页。
[13] 林恩·马古利斯、多里昂·萨根:《微观世界:微生物进化40亿年》(伦敦:亚伦和乌温出版社,1987年),第48页。
[14] 氨基酸是包含氨基群(NH2)与羧酸群(COOH)的简单的有机分子,也是依附于一个碳原子而数目不定的一群其他原子的组合;核苷也同样十分简单,由一个糖分子、一组磷酸盐(phosphate)群和四个碱基中的一个[包含氮(nitrogen)的化合物]所组成。长链将之链接在一起,核苷形成了两种主要类型的核酸(nucleic acids)脱氧核糖核酸和核糖核酸,由于它们最早是从细胞核(cell nuclei)中分离出来的,因此叫作核酸。脱氧核糖核酸和核糖核酸是一切生物遗传的关键。
[15] 早期生命领域的研究先驱马尔科姆·沃尔特(Malcolm Walter),曾在《探索火星生命》(悉尼:亚伦和乌温出版社,1999年)一书中对这些议题做过较好的论述。
[16] 达尔文语,转引自保罗·戴维斯:《第五奇迹》(哈蒙斯沃思:企鹅出版社,1999年),第54页。
[17] A. G. 凯恩斯——史密斯:《生命起源的七大线索》(剑桥:剑桥大学出版社,1985年)。
[18] 里夫斯、德·罗斯奈、科庞、西莫内:《宇宙、地球和人类的起源》,第92页(罗斯奈撰写的部分)。
[19] 关于原始细菌的这些发现,其详细讨论及意义,可参见戴维斯:《第五奇迹》,第7章。
[20] 卡伦·L.冯·达姆(Karen L. Von Damm):《失落城市的发现》,载《自然》,2001年7月12日,第127—128页。
[21] 比格尔·拉斯穆森(Birger Rasmussen):《32.35亿年前火山硫化物堆积中的纤维状微化石》,载《自然》2000年6月8日,第676—679页。在论文中,拉斯穆森宣布“嗜极生物”(extremophile)是至今为止所知最古老的化石,距今年代为32亿年,发现于澳大利亚的皮尔巴拉(Pilbara)。也许这些细菌靠“吃”化学物质为生,生活在海洋深处富含硫的火山口附近。在这以前,原始细菌最古老的遗迹只有5亿年。
[22] 关于进化孰先孰后,在弗里曼·戴森的《生命的起源》(剑桥:剑桥大学出版社,1999年)第2版中,对这个问题有比较清楚的论述。
[23] 切萨雷·埃米利亚尼:《科学指南:通过事实、数字和公式探索宇宙物理世界》第2版(纽约:约翰·威利出版社,1995年),第151页。弗雷德·霍伊尔曾有过一个著名的比喻,自行形成最简单细菌的概率,就像一堆废料在龙卷风的袭击下自行组成了一架波音747飞机。
[24] 关于这些深奥机制的具体描述,可参见考夫曼:《在宇宙的家庭中》,以及保罗·戴维斯:《宇宙蓝图》(伦敦:亚伦和乌温出版社,1989年)。
[25] 在约翰·梅纳德·史密斯和厄尔什·绍特马里(Eörs Szathmáry)所著的《生命的起源:从生命诞生到语言起源》(牛津:牛津大学出版社,1999年)第3章和第4章中,对“核糖核酸世界”有相关的描述。
[26] 戴森:《生命的起源》,第40页。
[27] 戴森:《生命的起源》,第3章。