
物理学家薛定谔。其著作《生命是什么?》启发了我投入基因领域。
我在芝加哥大学三年级时,迷上了基因。我原本想当博物学家,向往日后能离开自幼成长的芝加哥南区,到没有都市尘嚣的地方发展职业生涯。让我改变心意的,并不是某位难忘的老师,而是1944年出版的一本薄薄的小书《生命是什么?》(What Is Life?),作者是奥地利籍的波动力学之父薛定谔(Erwin Schrodinger)。这本书辑录了他前一年在都柏林的高等研究院(Institute for Ad-vanced Study)发表的数场演讲。这么伟大的物理学家竟会花时间写生物学的书,引起了我的兴趣。当时我和大多数的人都认为,化学与物理学才是“真正的”科学,而理论物理学更是科学翘楚。
以我为例,薛定谔触动了我的心弦,因为我对生命的本质也很感兴趣。当时仍有少数科学家认为,生命仰赖全能的上帝赋予生命力。不过如同我大多数的老师,我也鄙视生机论的观念。如果这种“生命力”是大自然运作的主宰,我们势必很难经由科学方法来了解生命。反之,一想到生命可能借由一本以密码写成的指令书而永续长存,就令我神往不已。什么样的分子密码能够复杂到足以传递众多的生命奇迹?又有什么样的分子秘密,让染色体在复制时都能复制出一模一样的密码?

薛定谔认为,我们可以从储存与传递生物信息的观点来思索生命。因此,染色体只是信息的携带者。由于每个细胞都要容纳这么多的信息,因此这些信息必须压缩成薛定谔所谓的“遗传密码脚本”(hereditary code-script),植入染色体的分子结构内。要了解生命,就必须辨识这些分子,破解它们的密码。他甚至臆测,了解生命(包括找到基因)说不定能让我们超越当时所知的物理定律。薛定谔这本著作的影响甚巨,日后许多在分子生物学这出大戏的序幕中成为要角的人物,包括我和克里克(他先前是物理学者)在内,都拜读过这本书,而且深受感动。
在薛定谔于都柏林演讲的年代,大多数的生物学家都认为,最终科学界会证明蛋白质是遗传指令的主要携带者。蛋白质是由20种不同的建构单元(氨基酸)所组成的分子链。由于氨基酸沿分子链排列的顺序可以说有无限多种,因此原则上蛋白质是有可能隐含造成生命如此多样的密码信息的。虽然DNA就位于染色体上,为世人所知也有75年之久,但当时并未将DNA视为密码脚本的可能携带者。1869年,在德国工作的瑞士生化学家米舍(Friedrich Miescher),从当地医院沾满脓的绷带上分离出一种物质,并称之为“核素”(nuclein),脓大多由具有细胞核的白血球构成(红血球没有细胞核),因此也具有包含DNA的染色体,米契尔等于在无意间发现了DNA的良好来源。稍后当他发现惟有在染色体里才找得到“核素”时,就知道自己有了重大发现。1893年,他写道:“遗传确保形态能世代延续,而这一切就隐藏在比化学分子还深的层次。它隐藏在结构化的原子群组内。因此,我支持化学遗传论。”
然而,数十年后,化学仍无法分析庞大复杂的DNA分子。一直到20世纪30年代,科学家才证明DNA是由四种不同的化学碱基所构成的长分子,即腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)与胞嘧啶(C)。不过在薛定谔发表演说的年代,科学界尚不明白DNA分子上的这些次单位(称为脱氧核糖核苷酸[deoxynucleotide])在化学上如何链接,也不知道DNA分子的四种化学碱基序列是否有差异。如果DNA真的是薛定谔所谓的密码脚本,那么这种分子应该有极多种不同的形式。不过在当时,一般仍认为整条DNA链有可能是由一个简单序列(例如AGTC)一再重复出现而构成的。
搜寻遗传物质:左图为显微镜下用DNA着色剂处理过的血球细胞。为使携氧能力达到最大,红血球没有细胞核,因此也没有DNA。但是在血流里搜寻入侵者的白血球有细胞核,内含染色体。
一直到1944年,DNA才成为遗传界的焦点,当时艾弗里(Oswald Avery)在纽约洛克菲勒研究所(Rockefeller Institute)的实验室发表报告说,肺炎病菌的外膜组成可以改变。这个结果出乎他和麦克劳德(Colin Macleod)与麦卡提(Maclyn McCarty)这两位资历较浅的同事意料。
艾弗里的研究小组花了10余年的时间,持续追踪英国卫生部科学家格里菲斯(Fred Griffith)在1928年视察到的奇特现象。格里菲斯对肺炎很感兴趣,潜心研究肺炎的致病菌——肺炎双球菌(Pneumococcus)。当时已知肺炎双球菌有两种形态,依照它们在显微镜下的外观而分为“平滑”(smooth)的S型与“粗糙”(rough)的R型。两者不仅外观不同,毒性也不同。将S型注入老鼠体内,几天内老鼠就会死去,但是注入R型的老鼠则依旧健康。后来发现S型的细胞有荚膜,可以防止老鼠的免疫系统认出它是入侵者。R型的细胞没有荚膜,因此会受到老鼠免疫系统的攻击。
格里菲斯从参与公共卫生的经验中得知,单一病人身上有时能分离出多种类型的菌株,因此他很好奇,不同类型的菌株在实验鼠身上会如何交互作用。后来他在一种组合上有了重大发现:当他将加热杀死的S型菌株(已变得无害)及正常的R型菌株(原本就无害)同时注入老鼠体内时,老鼠会死亡。两种无害的菌株在混合后,怎么可能变得致命呢?后来他分离出死老鼠身上的肺炎双球菌,发现里面有活的S型菌,线索于是出现。无害的R型菌似乎会从已死的S型菌取得不明物质;无论此物质为何,它显然会使R型菌在有加热致死的S型菌存在时,转型为活的杀手型S菌。格里菲斯从死老鼠身上培育出数代的S型菌,证实这种变化的确存在:这些细菌繁殖成S型菌,如同任何正常的S型菌株。注入老鼠体内的R型菌真的发生了“遗传”变化。
尽管这种转型现象似乎与当时的了解相悖,但格里菲断的观察结果起初并未在科学界激起太大的涟漪。部分原因在于格里菲斯非常注重隐私,厌恶大型聚会,鲜少参加科学会议,有一次还是在别人强迫下才发表演讲。那次他被同事架到出租车上,护送至演讲厅,然后以含糊单调的声调发表演说,谈的是他在微生物领域所作的艰涩研究,完全没有提及细菌转型。幸好,并非所有人都忽视格里菲斯的突破。
艾弗里对肺炎双球菌糖衣般的荚膜也很感兴趣。他复制格里菲斯的实验,试图分离出使R型菌变成S型菌的物质,找出它的特征。1944年,艾弗里、麦克劳德与麦卡提公布了他们的研究结果:他们以精心设计的一组实验明确地证实,DNA就是造成这祌变化的“转化因子”(transforming factor)。艾弗里及其研究小组用试管培养细菌,而不是用老鼠,因此更容易在加热致死的S菌细胞上,找出哪个化学物质是转型因子。他们有系统地一一破坏经过加热处理的S型菌的生化成分,看要摧毁哪一种成分,才能阻止转型发生。
首先他们使S型菌糖衣般的荚膜水解,但转型仍旧发生,这证明荚膜并非转化因子。接着他们使用两种酶(胰蛋白酶[trypsin]与胰凝乳蛋白酶[chymotrypsin])的混合制剂,这两种酶都会破坏蛋白质,结果S型菌的蛋白质几乎全遭破坏。出乎他们意料,转化仍然继续发生。他们又尝试一种会分解核糖核酸(ribonucleic acid, RNA)的核糖核酸酶(ribonuclease, RNase),但是转化再次发生。RNA也是一种核酸,与DNA类似,而且可能与蛋白质的合成有关。最后他们把目标锁定DNA,让从S型菌取出的萃取物接触会破坏DNA的脱氧核糖核酸酶(deoxyribonuclease, DNase),这次他们总算命中目标,R型菌不再转型为S型菌,转型因子就是DNA。
艾弗里、麦克劳德与麦卡提在1944年2月提交研究报告后,科学界反应不一,部分原因在于这项发现太过惊人。有许多遗传学家接受他们的结论,毕竟DNA在每个染色体上都找得到,它为什么不能是遗传物质?但是,也有许多生化学家对DNA分子是否复杂到能储存庞大的生物信息,表示存疑。他们仍旧认为,最终会证实同为染色体构成要素的蛋白质才是遗传物质。其实也难怪生化学家会这么想,因为基本上蛋白质有20个氨基酸字母可以编码庞大的信息,这要比只有4个核酸字母的DNA容易得多。跟艾弗里同在洛克菲勒研究所任职的蛋白质化学家墨斯基(Alfred Mirsky)更是激烈反对DNA是遗传物质的说法,不过那时艾弗里已不再活跃于科学界,洛克菲勒研究所强制他在65岁时退休。
艾弗里错失的,不仅是反击同事、为自己的研究成果辩护的机会,他也错失了获得诺贝尔奖的机会,身为DNA是转化因子的发现者,他其实很有获奖资格。由于诺贝尔奖委员会在各个奖项颁发50年后会公布记录,现在我们已经知道当时阻挡艾弗里获得候选资格的人,是瑞典籍的物理化学家哈马斯滕(Einar Hammarsten)。虽然哈马斯滕的声望主要奠基于他能萃取出质量绝佳的DNA样本,但他仍然相信基因是某类尚未辨识出来的蛋白质。事实上,即使在发现双螺旋后,一直到DNA转型的机制完全公布前,哈马斯滕仍坚持艾弗里不应获得诺贝尔奖。艾弗里于1955年过世,若是他再多活几年,肯定可以拿到诺贝尔奖。
1947年秋,我到印第安纳大学,计划以研究基因作为博士论文的题目,那时我们经常讨论艾弗里的论文。当时已没有人怀疑他实验结果的正确性,而洛克菲勒研究所也有更多的研究结果出炉,显示蛋白质不太可能是细菌转型过程中的遗传因子。至此,化学家终于把下一次重大突破的目标放在DNA上。英国剑桥精明干练的化学家托德(Alexander Todd)迎接挑战,开始鉴定连结DNA核苷酸的化学键。到了1951年初,他的实验室证实这些连结总是相同的,亦即DNA分子的骨干非常规则。同一时期,在奥地利出生,后来逃到美国,任职于哥伦比亚大学的内外科学院的查加夫(Erwin Chargaff)使用滤纸层析法(paper chromatography)这种新技术,萃取出多种脊椎动物与细菌的DNA样本,测量DNA内四种碱基的相对含量。他发现,有些物种的DNA以腺嘌呤与胸腺嘧啶居多,有些物种则是鸟嘌呤与胞嘧啶较多。因此,有可能任何两个DNA分子的组成都不同。
在印第安纳大学,我加入一小群以物理学家和化学家为主、具有远见的科学家之中,共同研究感染细菌的病毒“噬菌体”(bacteriophage,简称phage)的繁殖过程。后来我的博士论文指导老师,也就是在意大利受训的医师卢里亚(Salvador Luria),以及他在德国出生的理论物理学家好友德尔布吕克(MaxDelbrück),和美国理化学家赫尔希(Alfred Hershey)携手合作,成立了噬菌体研究小组(the Phage Group)。在第二次世界大战期间,尽管犹太裔的卢里亚是被迫离开法国投奔纽约,德尔布吕克因为反对纳粹主义而逃离德国,但他们仍被视为敌侨,没有资格和美国科学界共同为战争效力。虽然遭到排挤,但他们还是在各自的大学实验室里努力工作(卢里亚在印第安纳大学,德尔布吕克则在范德比尔特大学),并且接连几年夏季都到冷泉港实验室合作进行噬菌体实验。1943年,他们与才华洋溢但沉默寡言的赫尔希合作,当时赫尔希正在圣路易的华盛顿大学研究噬菌体。
噬菌体小组认为,如同所有的病毒,噬菌体其实就是赤裸裸的基因(病毒就只是一个蛋白质外鞘包着核酸),并依据这个想法来规划研究计划。这个概念是想像力丰富的美国遗传学家穆勒(Herman J. Muller)于1922年首先提出的,三年后,他证实X光会引起突变。但是直到1946年,穆勒来印第安纳大学任教不久之后,才获得了迟来的诺贝尔奖。事实上,正是因为他在印大,我才会到这里就读。穆勒是在摩根的手下展开他的职业生涯的,遗传学在20世纪前半叶的发展过程,没人比他更清楚,我在第一个学期时就对他的讲学非常着迷。然而,他在果蝇方面的研究,对我而言似乎属于过去,而非未来,因此我只短暂考虑过请他指导我的论文。后来我选择卢里亚的噬菌体实验,这个实验做起来比果蝇快得多:噬菌体的遗传杂交(genetic cross)子代在隔天就可以进行分析。
为了完成博士论文研究,我在卢里亚的要求下跟随他的步履,研究X光如何杀死噬菌体粒子。刚开始时,我希望能证明病毒死亡是因为噬菌体的DNA遭到破坏。但最后我不得不承认,我的实验方法在化学上永远无法获得确切的答案,只能得到生物学上的结论。虽然噬菌体的确是裸露的基因,但我知道噬菌体研究小组所要的深奥答案,惟有通过高深的化学才找得到。
DNA不再只是一个笼统的缩写名词,我们必须彻底解开它的分子结构和所有的化学细节才行。
完成论文后,我发现自己别无选择,只能到可以让我研究DNA化学组成的实验室。然而,不幸的是,由于我几乎毫无理论化学的基础,实在不够格转战任何以有机化学或物理化学来进行艰难实验的实验室。后来在1950年秋,我拿到博士后研究奖学金,到生化学家开尔卡(Herman Kalckar)在哥本哈根的实验室作研究。当时他正在研究构成DNA的小分子的合成作用,但我很快就发现,他的生化方法永远无法解开基因的本质。在他的实验室多待一天,就会晚一天了解DNA如何携带遗传信息。
不过,我在哥本哈根的那一年仍然获益良多。为了避开丹麦寒冷的春天,我在四五月间前往意大利那不勒斯动物研究所。在那里的最后一周,我参加了一场小型研讨会,主题是以X光衍射法(X-ray diffraction)决定分子的三维结构。X光衍射法可以研究任何能够形成晶体的分子的原子结构。X光在轰击晶体后,会在撞到原子时弹开而散射。从X光的散射图形可以获得有关分子结构的信息。但是只靠X光,尚不足以解决结构的问题,还需要所谓“相分配”(phase assignment)的额外信息,来处理分子的波性质(wave properties)。要解决“相”的问题并不容易,当时只有胆量最大的科学家愿意面对这种挑战。以衍射法成功研究的对象,大多是比较简单的分子。
原本我对这场研讨会的期望不高,因为我认为要解开蛋白质或DNA的三维结构,起码还要10年光景。从早期令人失望的X光照片看来,要通过X光来解开DNA的秘密,尤其不可能。会有这种结果也是很自然的,因为当时大家都预期,每个分子的DNA序列应该不尽相同。在表面构造不规则的情况下,DNA细长的分子链势必不可能按照规律重复的模式整齐地排列,X光分析自然也无法成功。
因此,当我听到来自伦敦国王学院(King's College)生物物理实验室的英国人威尔金斯(Maurice Wilkins)在最后发表有关DNA的演讲时,不禁惊喜交加。34岁的威尔金斯是物理学家,战时曾参与制造原子弹的曼哈顿计划(Manhattan Project)。对他和许多参与这个计划的科学家而言,原子弹投到广岛和长崎,应该是他们研究工作的最高成就,结果这却造成他们的理想破灭。威尔金斯曾考虑完全放弃科学,到巴黎去当画家,但生物学引起了他的兴趣——他也读过薛定谔的书。当时他正设法用X光衍射法解开DNA的秘密。
威尔金斯在演讲中展示一张最近拍到的X光衍射图,上面有许多明确的反射影像,显示它是高度规则的结晶体。由此可以推论出,DNA必定具有规则的结构,只要能解开此结构,就可以揭露基因的本质。我立即开始幻想自己搬到伦敦去,协助威尔金斯找出这个结构。演讲过后,我去找他谈话,但却一无所获。他只对我表示,未来还有更多艰辛的工作要做。
就在我连续碰壁时,加州理工学院享誉国际的化学家泡令(Linus Pauling)宣布获得了重大的成就:他发现蛋白质里氨基酸链(称为多肽[polypeptide])的排列结构,并且将这个结构取名为α螺旋(alpha helix)。这个突破会由泡令获得其实并不意外,他是科学界的超级巨星。他所著的《化学键的本质》(The Nature of the Chemical Bond)奠立了现代化学的基础,被当时的化学家奉为圣经。泡令非常早熟,他在俄勒冈州长大,父亲是位药剂师。他9岁时,父亲曾写信给《俄勒冈人》报,希望对方能提供他那好学不倦的孩子可以阅读的书籍,还说他儿子已经读完圣经和达尔文的《物种起源》。泡令的父亲不幸早逝,家中经济陷入困境,但这位前途无量的年轻人仍然完成了学业,相当难能可贵。
我一回到哥本哈根,立即拜读了泡令有关α螺旋的研究。令人惊讶的是,他并非根据X光衍射的实验数据推论出模型,而是根据身为结构化学家的丰富经验,大胆推论哪种类型的螺旋结构最符合多肽链的化学特性。泡令制作蛋白质分子不同部分的比例模型,找出可能的三维结构。他将问题简化成一种三维拼图游戏,既简单又聪明。
α螺旋相当美丽,但现在的问题在于它是否正确。短短一星期后,我得到了答案。发明X光晶体学(X-ray chrystailography)的1915年诺贝尔物理学奖得主布喇格爵士(Sir Lawrence Bragg)来到哥本哈根,兴奋地宣布,比他资浅的同事、奥地利籍化学家佩鲁茨(Max Perutz)巧妙地用合成多肽证实了泡令的α螺旋是正确的。对布拉格的卡文迪什实验室而言,这是个苦乐参半的胜利,因为前一年他们在论文中列举多肽链可能具有的螺旋形态时,完全不得要领。
那时,卢里亚尝试替我安排卡文迪什实验室的研究职位。卡文迪什位于剑桥大学,是科学界最著名的实验室,卢瑟福(Ernest Rutherford)就是在这里首先描述出原子的结构。当时那是布喇格的研究领域,而我则被安排跟英国化学家肯德鲁(John Kendrew)实习,他的兴趣是找出肌红素(myoglobin)这种蛋白质的三维结构。卢里亚建议我尽快前往卡文迪什实验室,因为肯德鲁那时在美国,佩鲁茨会审核我的资格。肯德鲁与佩鲁茨早先曾一起建立医学研究委员会(Medical Research Council, MRC)作为生物系统结构研究的单位。

布拉格(左)与拿着α螺旋模型的泡令合影。
一个月后在剑桥,佩鲁茨向我保证,我很快就能精通必要的X光衍射理论,应该也很快就能融入他们人数不多的研究单位。我松了一口气,因为他并没有因为我的生物学背景而拒绝我。布喇格也没有,他还从办公室下来看了我一下。
我在10月初抵达剑桥医学研究委员会的研究单位,那年我23岁。我和35岁的前物理学家克里克共用生化研究室,他在战时曾替英国海军研究磁性水雷。战争结束后,克里克原本计划留在军方的研究机构,但在拜读了薛定谔的《生命是什么?》后,决定朝生物学发展。当时他在卡文迪什实验室以研究蛋白质的三维结构为博士论文。
我和克里克相遇的那个早晨,他听说我来剑桥是为了大量学习有关晶体学的知识以便破解DNA的结构,大为高兴。不久之后,我就请克里克谈谈他对使用泡令的模型建构法来破解DNA结构的看法。我们是不是还得做许多年的衍射实验,才能实际下手去建构模型?为了加快我们研究DNA结构的速度,克里克邀请自战后就认识的朋友威尔金斯,在星期天从伦敦过来共进午餐,这样我们就能得知自从那不勒斯的演讲后,威尔金斯还有哪些进展。

威尔金斯,摄于伦敦国王学院的实验室。
威尔金斯表示,他认为DNA的结构是螺旋状,由数条链接的核苷酸互相缠绕而成,而惟一尚待解决的问题在于链的数目。当时威尔金斯根据他测量的DNA纤维密度,认为应该有三条核苷酸链。他迫切地想着手建造模型,却碰上了一个障碍:刚加入国王学院生物物理学研究单位的富兰克林(Rosalind Franklin)。
富兰克林才从法国回来,她在巴黎用X光晶体学的技术研究了4年的石墨。她接受聘任,加入国王学院的DNA计划时,威尔金斯刚好不在。不幸的是,后来证明这两人根本合不来。富兰克林的个性率直,重视数据,威尔金斯则拘谨而勇于猜想,他们注定无法合作。在威尔金斯接受我们的午餐邀约前不久,他们两人才大吵一次,富兰克林坚持在她搜集更多衍射数据前,不能着手建立模型。他们俩显然无法沟通,而在富兰克林于11月初举办实验室研讨会之前,威尔金斯无从得知她的进展。不过若我们想参加这次研讨会,威尔金斯很乐意邀我和克里克前往。后来克里克因故未能参加研讨会,由我独自前往,稍后再把我认为与DNA晶体有关的重要讯息告诉他。我特别根据记忆,描述了富兰克林关于晶体重复与含水量的测量值。克里克听了之后,开始在纸上绘制螺旋网格,并说就连我这种先前以赏鸟为业的人也可以应用他和科克兰(Bill Cochran)及凡德(Vladimir Vand)提出的新螺旋X光理论,准确预测我们即将建造的分子模型有哪些衍射图。

富兰克林在假期时热爱登山运动,摄于欧洲阿尔卑斯山。
31岁的富兰克林是出身剑桥的物理化学家,她是个对自己的专业执着得不得了的科学家,在29岁生日时,她只要求订阅自己所属领域的技术期刊《晶体学报》(Acta Crystallogrsphica)作为生日礼物。她讲求逻辑和精确,对于欠缺这些特质的人没有什么耐性。她不习惯快速发表措辞强烈的看法。一度将她的博士论文指导教授,未来的诺贝尔奖得主诺里什(Ronald Norrish)描述为“愚蠢、固执、奸诈、态度恶劣、专制”。在实验室外,她是果决勇敢的登山家,来自伦敦的上流社会,相较于大多数的科学家,她属于高尚的社交界。在工作台辛苦一整天后,她偶尔会脱下实验室的外套,换上优雅的晚礼服,消失在夜色中。
克里克对重要问题的错综复杂总是非常着迷。小时候,他老爱问问题,被问腻的双亲只好买一套儿童百科全书给他,希望能满足他的好奇心,结果这反而让他没有安全感,他告诉母亲,他怕长大时,所有的事物都已被人发现,而他将无事可做。母亲向他保证,日后一定还会有一两件事等着他发现。事后证明她说的很准。
克里克很擅长说话,无论是哪种聚会,他总是众人注意的焦点。在卡文迪什实验室的走廊上,总是可以听到他爽朗的笑声。他是医学研究委员会研究单位的专任理论家,每个月至少会提出一个新构想,而且只要有人愿意听,他总是很乐意花许多时间仔细解释。

克里克与卡文迪什实验室的X光管。
我们一回到剑桥,我就安排卡文迪什的机械部门建造磷的原子模型,以便用于建造DNA里磷酸糖骨干的片段。等这些模型做好后,我们开始测试骨干在DNA分子中央彼此缠绕的不同方法。它们规则重复的原子结构,应该会让原子形成一致且重复的构造。我们听从威尔金斯的直觉,把重点放在三链模型上。当其中一个模型看似很有可能是答案时,克里克打电话给威尔金斯,宣称我们可能找到了DNA的模型。
第二天,威尔金斯与富兰克林一起来访,査看我们的成果。在这意外出现的竞争威胁下,他们俩为了共同目标而难得地暂时合作。富兰克林立即挑出我们在基本概念上的错误。我记得她在报告时指出:DNA晶体几乎不含水。其实是我自己弄错了。由于才刚开始学晶体学,我把晶胞(unit cell)与不对称单位(asymmetric unit)这两个术语搞混了。其实,DNA晶体富含水分。因此,富兰克林指出,光是要容纳她在晶体内观察到的水分子,骨干就得在分子外面,而不是像我们所做的在分子中央。
11月的那一天实在不幸,让未来蒙上浓浓阴影。富兰克林更加坚定地反对建造模型,她打算继续做实验,不玩看似小孩玩具的原子模型。更惨的是,布拉格也说话了,叫我和克里克不要再尝试建造DNA模型,后来还进一步决定,DNA研究应交由国王学院实验室来做,剑桥只需继续研究蛋白质即可。两家同样由医学研究委员会赞助的实验室居然彼此竞争,实在没有道理。在无计可施下,我和克里克不情愿地暂时罢手。
在此时退出DNA研究,实在不是时候。泡令已写信给威尔金斯,请他提供一份DNA晶体的衍射图。虽然威尔金斯拒绝,表示自己需要更多的时间来解读,但泡令其实也不见得要依赖国王学院的资料。如果他愿意的话,大可在加州理工学院自行研究X光衍射。
来年春天,我不再研究DNA,反而继续作那些战前的研究,用卡文迪什实验室强大的新X光束,研究铅笔状的烟草花叶病毒(tobacco mosaic virus)。
这个实验的工作量很少,我有许多时间游走剑桥众多的图书馆。我在动物学系看到查加夫的论文,他发现在DNA中,腺嘌呤和胸腺嘧啶的数量大致相同,而鸟嘌呤则与胞嘧啶的数量差不多。在听到这一比一的比例后,克里克想到,在DNA复制时,腺嘌呤和胸腺嘧啶是否互相吸引,而胞嘧啶与鸟嘌呤之间是否也存有类似的吸引力。若是如此,DNA“亲代”链上的碱基序列(例如ATGC)应该会与“子代”链上的互补(亦即TACG)。
这些原本都只是空想,直到1952年夏天,査加夫在前往巴黎参加国际生化会议时途经剑桥为止。我和克里克认为不需要了解4种碱基的化学结构,但査加夫对这看法颇不赞同。又听到我们说,如有必要的话,可以到教科书里查它们的结构时,他更是不悦。我只希望能证实查加夫的数据与DNA结构并不相关。不过克里克却兴致勃勃地要做一些实验,寻找腺嘌呤与胸腺嘧啶(或鸟嘌呤与胞嘧啶)在溶液中混合时,可能会形成的分子“三明治”。但是,他的实验没获得任何结果。
泡令跟査加夫一样,也参加了国际生化会议,那时的大新闻是噬菌体研究小组的最新结果。冷泉港的赫尔希与蔡斯(Martha Chase)才刚证实了艾弗里的转化因子:DNA就是遗传物质!赫尔希和蔡斯证明,进入细菌细胞的只有噬菌体病毒的DNA,它的蛋白质鞘(protein coat)留在外面。看来如果我们想揭开基因的本质,势必得了解DNA的分子。在赫尔希与蔡斯成为大家的话题之后,我确定泡令也会将他的才华与化学知识,全力投注在解决DNA的问题上。
早在1953年,泡令就发表过描述DNA结构的论文。我急切地拜读了大作,发现他提出的是三链模型,以磷酸糖的骨干形成稠密的中央核心。乍看之下,它跟我们在15个月前所做的拙劣模型类似。但是,泡令没有采用带正电的原子(例如Mg2)来稳定带负电的骨干,而是采取非正统的做法,以氢链来连接磷酸盐。不过,看在身为生物学家的我眼中,这种氢键所需的极酸状态从不曾见于细胞内。我发疯似的冲到托德在附近的有机化学实验室,立刻就证实了我的看法:不可能的事居然发生了!全世界最优秀或至少最著名的化学家竟然弄错了化学基本原理。实际上,泡令等于把DNA里代表酸的缩写A除掉了。我们研究的对象是脱氧核糖核酸,但是他所提出的结构甚至不属于酸类。
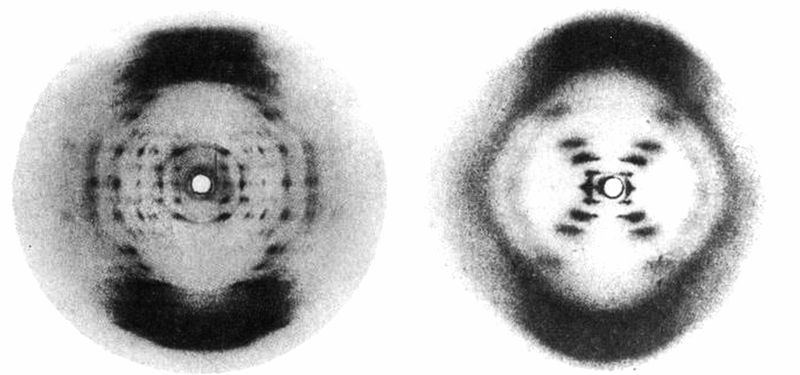
A型DNA与B型DNA的X光照片,分别来自威尔金斯与富兰克林。分子结构的差异是由每个DNA分子水含量的差异所造成。
我立刻把论文带到伦敦,告诉威尔金斯与富兰克林,他们仍有成功机会。但深信DNA并非螺旋的富兰克林甚至不想看这篇文章,以免受到泡令的螺旋观念影响,连在我提出克里克的螺旋论点后也没改变想法。倒是威尔金斯对我带来的消息很感兴趣;他现在更确定DNA是螺旋。为了证明这一点,他拿出一张6个月前,由富兰克林的研究生葛斯林(Raymond Gosling)用X光拍下的照片,即所谓的B型DNA。在那之前,我甚至不知道有B型的存在。
富兰克林不理会这张照片,把注意力集中在A型DNA上,因为她认为研究A型比较可能获得有用的资料。B型DNA的X光图是一个清晰的十字。既然克里克和其他人早已推论出,这类的反射图案是由螺旋所造成,这项证据清楚说明DNA必定是螺旋状!事实上,尽管富兰克林持保留态度,但这个发现并不出人意表。几何学本身就显示,螺旋结构是一长串重复的单元(例如DNA的核苷酸)最合理的排列方式。不过我们仍不知道这个螺旋的外观,也不知道它含有多少链。
现在终于到了我们继续建构DNA螺旋模型的时候。泡令肯定不久就会发现,他的精心杰作出现了谬误。我敦促威尔金斯不要再浪费时间,但是他想等到富兰克林在该年春天稍后到另一家实验室工作后才开始。她选择离去,避开在国王学院的不愉快。在离开前,她奉命停止对DNA作更进一步的研究,并将许多衍射照片交给威尔金斯。
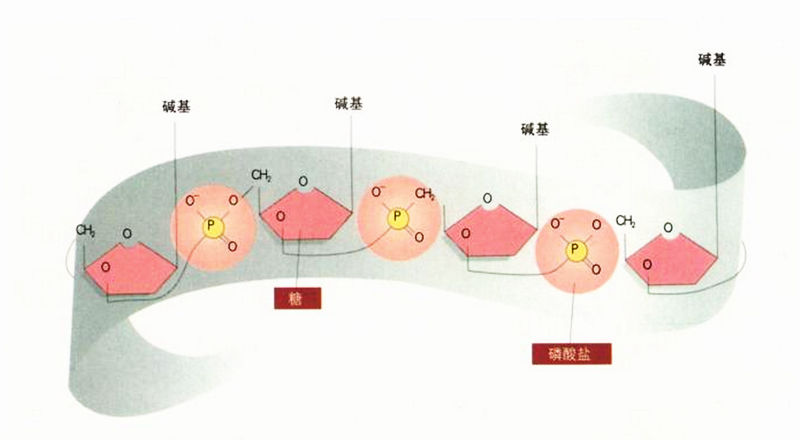
DNA的化学骨干
当我回到剑桥,向布拉格报告B型DNA的消息之后,他认为不应再禁止我和克里克研究DNA,而且很希望DNA的结构能由大西洋这一岸破解。于是我们再度着手建构模型,设法把已知的DNA基本成分凑成螺旋结构。这些基本成分就是分子骨干以及4个不同的碱基(腺嘌呤、胸腺嘧啶、鸟嘌呤与胞嘧啶)。我委托卡文迪什的工厂替我们做一套锡制的碱基模型,但是他们的制造速度对我来说不够快,最后我只得拿硬纸板来剪出粗略的模型。
此时我已经发现,DNA的密度测量证据比较倾向于双链,而非三链的模型结构。因此,我决定寻找可能的双螺旋体。身为生物学家,我偏好遗传分子是由两个,而非三个成分组成的概念。毕竟,染色体就像细胞一样,数量是以复制成两倍而非三倍的方式增加。
我知道先前将骨干置于里面,而碱基悬挂在外的模型是错误的。诺丁汉大学提出的化学证据显示,碱基必须由氢键彼此连结,但这项证据一直被我忽略。如果碱基位于分子中央的话,就只能按照X光衍射数据所显示的规律方式形成这种键。但是它们怎么会成双作对呢?在错误的核酸化学教科书误导下,我连续两个星期毫无进展。幸好2月27日加州理工学院的理论化学家多纳休(Jerry Donahue)到卡文迪什访问,他指出教科书的错误。于是我改变了氢原子在硬纸板分子模型上的位置。
隔天早晨,1953年2月28日,DNA模型的重要特征全都各就各位。它的两条链由腺嘌呤-胸腺嘧啶,以及鸟嘌呤-胞嘧啶这两对碱基对之间的强氢键连在一起。克里克一年前根据查加夫的研究所得到的推论,真的是正确的。
腺嘌呤的确与胸腺嘧啶连结,而鸟嘌呤也与胞嘧啶连结,但是它们并非通过平坦的表面形成分子三明治。当克里克抵达时,他很快就了解了状况,并且认同我的碱基配对(base-pairing)方式。而且他当下就发现,这会造成双螺旋的双股以相反方向连接。
这真是令人难忘的时刻,我们觉得这次肯定对了。这么简单美丽的构造,绝对错不了。最令我们兴奋的是碱基序列沿着双链排列的互补特性,只要知道一条链上的序列(碱基的顺序),自然就能推知另一条链上的序列。这显然是细胞分裂前染色体在进行复制时,基因的遗传讯息能精准重现的原因。
DNA分子会像拉链一样“拉开”,形成独立的两股。每一股都可以作为新股合成时的模板,于是一条双螺旋就变成了两条。
在《生命是什么?》中,薛定谔提议,生命的语言也许就像摩斯密码,是一系列的点与线。这个讲法倒是蛮接近事实的。DNA的语言是由A,T,G与C构成的线性序列。就像我们在誊写书籍时,偶尔也会打错字一样,所有的A,T,G,C在沿染色体复制时,也会出现极少量的错误。这些错误就是遗传学家近50年来所一直探讨的突变。在英文中,将i变成a,Jim就会变成Jam,而在DNA中,将T变为C,ATG就变成了ACG。
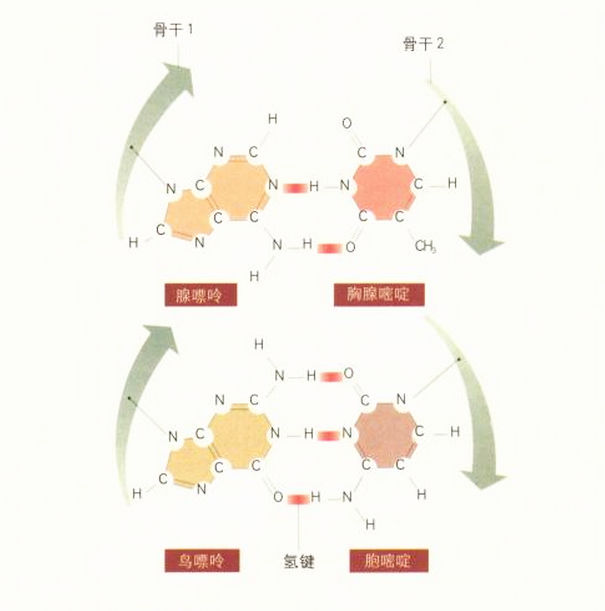
让一切运作的原理:碱基由氢键连结的互补配对。
无论从化学或生物学的观点来看,双螺旋都很合理。我们现在无须担忧薛定谔所说,要了解遗传密码如何复制有可能需要新的物理定律,事实上,基因的组成与其他的化学作用并没有两样。那天稍后,在紧邻卡文迪什实验室的鹰吧吃午餐时,向来爱说话的克里克忍不住告诉大家我们刚发现了“生命的奥秘”。我虽同样激动,但宁可等到做出漂亮的三维模型时才炫耀。
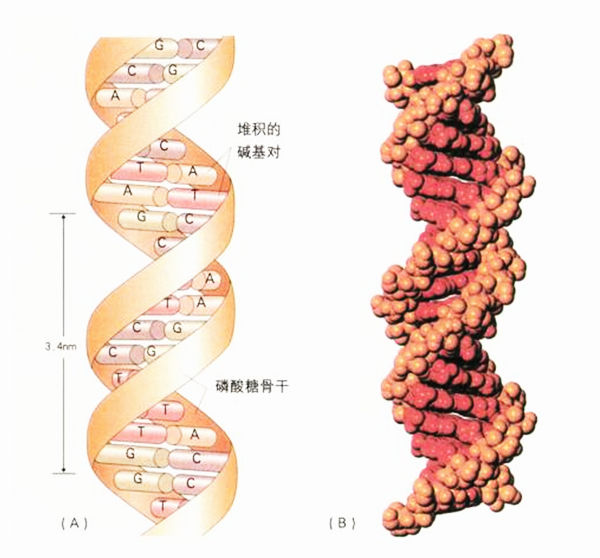
碱基与骨干就位,形成双螺旋:(A)将DNA的两股连结在一起的碱基配对系统;(B)将分子的原子细节按比例呈现的“空间填充”模型。
化学家托德是最早看到我们这个模型的人之一。基因的本质如此简单,让他非常惊喜。然而,稍后他必然曾扪心自问,为什么自己的实验室在建立DNA链的一般化学结构后,未能进一步研究这些链在三维空间的组成方式,反而让由生物学家和物理学家组成的双人组找出这种分子的本质,这两人对大学程度的化学甚至都不是那么了解。然而话说回来,这正是我们成功的关键,至少是部分关键:我和克里克之所以能率先获得双螺旋的结论,正是因为当时大多数的化学家认为,DNA的分子太大,无法用化学分析来了解。
同时,惟一两位具有远见、知道要寻找DNA三维结构的化学家,又犯了策略上的错误:富兰克林不愿建构模型,泡令则忽略了有关DNA的现有文献,特别是查加夫所发表的DNA碱基组成数据。1952年巴黎的生化会议后,泡令和查加夫还曾同船越过大西洋,两人却不投缘。泡令习惯于自己总是对的,而且相信自己可以用基本原理来解开任何化学问题。他的自信通常很有道理。冷战期间,他是批评美国核武器发展计划的主要人士,有一次在发表演说后,FBI警员质问他怎么知道原子弹含有多少钸?泡令傲然答道:“没人告诉我,我自己想出来的。”在其后的几个月,克里克和我(虽然我没那么热衷)乐得把我们的模型拿给川流不息、充满好奇的科学家们看。然而,剑桥的生化学家并未邀请我们到生化大楼发表正式演说。他们戏称这是WC,拿我们的名字缩写开双关语的玩笑。我们没做实验就找到双螺旋,令他们恼怒。
我们在4月初将发现双螺旋的报告交给《自然》(Nature)杂志,并于3个星期后,也就是1953年4月25日刊出。同期还有两篇由富兰克林与威尔金斯执笔、篇幅较长的论文,两篇文章都认为我们的模型大致正确。6月,我在冷泉港实验室的病毒研讨会上,首次就我们的模型提出报告。德尔布吕克亲自出马确保了我在最后一刻受邀发表演说。我带了在卡文迪什制造的三维模型,参加这场精英荟萃的会议,我使用的腺嘌呤-胸腺嘧啶碱基对模型是红色,而鸟嘌呤-胞嘧啶碱基对则是绿色。

小而美:沃森和克里克在《自然》杂志发表的短文,宣布他们发现了DNA的双螺旋结构,篇幅只有一页。同期也刊载了富兰克林与威尔金斯较长的文章。
同样受到薛定谔那本著作感召的前物理学家本泽(Seymour Benzer)也出席了这场盛会。他立刻了解到我们的突破性发现,对于他的病毒突变研究具有重要意义。他发现现在他能够以“摩根的孩子们”在40年前研究果蝇染色体的方式,来研究一小段的噬菌体DNA:他可以在基因图上标出突变位置,就好像当年研究果蝇的先驱们在染色体上标出基因的位置。本泽跟摩根一样,也得靠重组作用来产生新的遗传组合。不过摩根可以利用现成的重组机制,即果蝇性细胞的产物。本泽则得用两种不同的噬菌体同时感染一个的细菌宿主细胞,借此引发重组机制。这两种不同的噬菌体在重要区段有一个或多个突变的差异。在细菌细胞里,重组作用(分子片段的交换)有时会发生在不同的病毒DNA分子之间,产生新的突变置换,即所谓的重组体(recombinant)。在普渡大学(Purdue University)的实验室里,本泽短短一年内就有惊人的丰富成果,他制作了噬菌体rⅡ基因的图谱,显示出一连串的突变(遗传脚本上的所有错误)在病毒DNA上的线状排列方式。这种“生命语言”既简单又呈直线形状,就像书页上的一行文字。对于我在冷泉港所发表有关双螺旋的演讲,匈牙利物理学家齐拉特(Leo Szilard)的反应跟学术比较无关。他问我:“你能申请专利吗?”有一阵子,齐拉特的主要收入来源是他和爱因斯坦共享的一项专利,后来他还试图和费米(Enrico Fermi)一起申请1942年他们在芝加哥大学所建造的核子反应炉专利,但并未成功。不过当时和现在一样,专利只授予有用的发明,当时没有人想到DNA有什么实际用途。于是,齐拉特建议,我们应该申请版权。
解开双螺旋:1953年6月沃森在冷泉港实验室发表演说。
然而,在双螺旋的拼图中,还是少了一块:对于DNA“像拉链一样拉开”的复制方式,我们还需要以实验来证实。德尔布吕克就不相信我们的概念,他喜欢双螺旋的模型,但是担心把双螺旋拉开,会产生可怕的打结情况。5年后,泡令以前的学生梅索森(Matt Meselson)及同样聪明的研究噬菌体的年轻学者史塔尔(Frank Stahl),发表了一个简单明了的实验结果,一举扫除了这类疑虑。
1954年夏天,这两人在马萨诸塞州伍兹霍尔(Woods Hole)的海洋生物实验室结识,当时我在那里讲学。喝了不少马丁尼之后,两人决定他们应该搭挡研究科学。他们的合作结果赢得了“生物学上最完美的实验”之称。
他们使用离心技术,按照重量的些微差异来分离分子。在离心旋转后,较重的分子会比较轻的分子落到更接近试管底部之处。
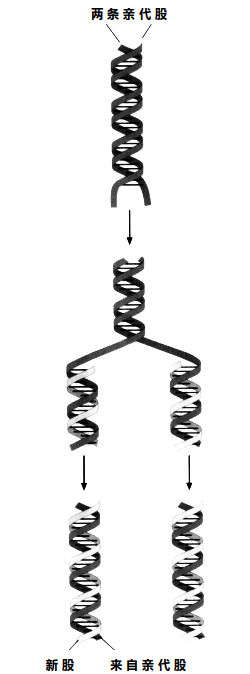
DNA复制:双螺旋先拉开,再各复制一股。
氮原子(N)是DNA的成分之一,而且原本就有两种不同的形态,一种较轻,另一种较重,因此梅索森和史塔尔能借由标注DNA片段,追踪DNA在细菌里的复制过程。起初所有的细菌都在含有重氮的培养基里培养,让重氮进入DNA的双股上。然后他们从这个培养菌中取出样本,转移至仅含轻氮的培养基,确保下一次DNA在复制时,只会用到轻氮。
如果DNA的复制如同我和克里克的预测,是将双螺旋拉开,然后各复制一股,那么实验所制造出的两个“子代”DNA分子将会是混种,每个分子都包括一个重氮股(来自“亲代”分子的模板股),以及一个轻氮股(用培养基制造出来的新股)。梅索森与史塔尔的离心处理程序完全证实了这种预测。他们发现离心试管分离出三个明显的区段,分别是重—重、轻—轻,以及介于两者之间、先重后轻的样本。DNA的复制方式跟我们的模型所预测的完全相符。
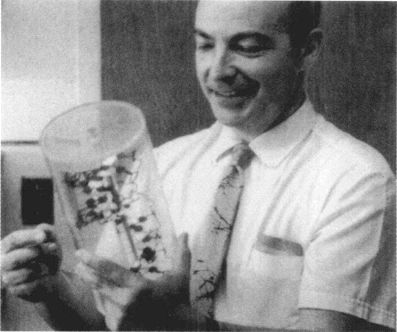
大约同一时间,酶学专家科恩伯格(Arthur Kornberg)位于圣路易市华盛顿大学的实验室也在分析DNA复制过程的生化细节。科恩伯格发展出一种新的合成DNA的无细胞系统,从而发现了一种称做DNA聚合肽(polymerase)的酶,这种酶连接构成DNA的不同小单元,形成DNA骨干的化学链。科恩伯格发现DNA酶合成机制,是惊人的重要大事,因此在这些实验完成后不到两年,他就于1959年获得了诺贝尔生理医学奖的殊荣。在公布得奖后,科恩伯格拿着我在1953年带到冷泉港实验室的双螺旋模型复制品摄影留念。
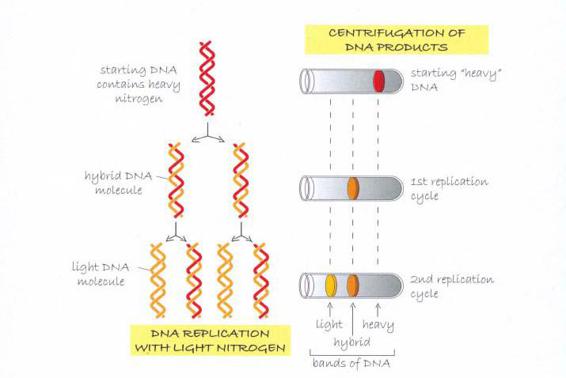
梅索森和史塔尔用细菌所做的实验。
直到1962年,克里克、威尔金斯和我才获得诺贝尔生理医学奖。在此4年前,富兰克林已经因卵巢癌而不幸早逝,享年37岁。那时克里克已和她成为亲密的同事与真正的好友。富兰克林动了两次手术,但都未能遏阻癌细胞蔓延,之后她曾回到剑桥,在克里克和他太太欧蒂莉(Odile)的照顾下疗养。
科恩伯格赢得诺贝尔奖,他手中拿着双螺旋模型的复制品。
诺贝尔委员会向来不曾将单一奖项颁给超过三个人,倘若当时富兰克林还在人世,他们势必得面对要将这个奖颁给她或威尔金斯的问题。瑞典人可能会授予他们诺贝尔化学奖以解决这个难题。最后,化学奖颁给了佩鲁茨和肯德鲁,他们分别发现了血红素与肌红素的三维结构。

梅索森站在超离心机旁,这是“生物学上最完美的实验”所使用的重要设备。
双螺旋的发现敲响了生机论的丧钟。认真的科学家,甚至有宗教信仰的科学家都已发现,要对生命有完整的了解,不需要寻找新的自然定律。生命不过就是物理与化学——尽管是极为精密复杂的物理与化学。接下来的工作是要找出,生命如何上演出隐藏在DNA中的密码脚本。细胞的分子机器如何读取DNA分子携带的讯息?下一章将介绍,极度复杂的读取机制如何引导我们深入了解生命的形成。