在本书的剩余部分我们将主要关注一个物种的历史:这个物种便是智人。之所以缩小我们的关注点,原因有二:首先,我们——本书的作者和读者——均属于这一物种,若想了解我们自己,就必须掌握智人的历史;其次,从某些异常巨大的尺度看,我们这一物种的历史具有相当的重要性,这一点虽然不太明显,却也并非褊狭之见。
当我们试图解释人类的出现时,我们再次面对关于起源的悖论。全新的事物是怎么出现的呢?我们是动物,和其他生物体一样依照达尔文的理论进化,我们和相近的亲缘物种,比如其他人科动物(大猿)有明显的相似之处,然而我们又与即使最相近的亲缘物种有着根本的不同。不知何故,我们这一物种的进化超越了达尔文的规律。正是这个原因,与其他大型生物体相比,我们对地球产生的影响要大得多。
那么,我们该如何解释是什么把我们与其他动物联系在一起,又是什么把我们与它们区别开来呢?
人类的历史:一个新层次的复杂性
前文我们已讲述过类似的跃迁。人类的历史标志着一个新层次的复杂性的出现是突如其来而不可预计的,就像最早的恒星、地球生命或是多细胞生物体的出现那样。我们看到,复杂实体比简单实体更加罕见,它们更加脆弱,而且由于不得不在熵的下行电梯上往上攀爬得更快一些(参见附录二),所以它们不得不获取更为密集的能量流。我们还看到,随着那些多少相互独立的实体合并为新的更大的结构,创造出了一种新形式的相互依赖,这种方式导致了更大复杂性的跃迁。最后,我们还看到,随着新层次复杂性的出现,它们似乎按照某种新的规则(“突现属性”,用复杂理论的术语来说)而发生作用。
人类的历史同样标志着地球上一种新层次复杂性的出现。[1]在较早的跃迁中,人类的历史将曾经独立的实体联结成为更大的相互依存的范型,这个过程是与造成深刻变化后果的大量能量流密切相关的。以21世纪的观点来看,我们能够估算其中的一些变化。人类由于集体行动而学会了掌握迅速增长的大量能量流。尽管这些变化的重大意义直到最近两个世纪里才显现出来,但是它们的根源可以追溯到旧石器时代。[2]
表6.1说明了人类怎样从环境中汲取远远超过他们生存和繁殖所需的能量。他们显现了一种全新的“生态创新”能力。从人类早期历史开始,如用火之类的技能提高了每人所能分配到的能量数量。在过去1万年里,农业提高了人类在一定范围内的食物能量,而过去6000年中大型食草动物的驯化则提高了牵引动力的能量。在过去两个世纪里,使用矿物燃料使人均分配到的能量成倍增长。随着人口总数大致从旧石器时代的几十万发展到1万年前的几百万,直到今天的超过60亿(参见表6.1),我们人类这个物种所掌控的能量,其数量至少增加了5万倍。对于一个物种所能支配的能量的数量来说,这是一个令人惊愕的数字,这也有助于我们解释为什么人类对于整个生物圈能够产生如此巨大的影响。要想衡量这种影响的一个有力方法就是估算一下日光所提供给生物圈的能量有多少是被人类所吸收和利用的。净初级生产力(NPP)是指从日光中吸收的能量,通过光合作用进入食物链而转化为植物。然后,它们喂养了许多其他生物体。这样,净初级生产力能够用来粗略地衡量生物圈的能量“收入”。现代的计算结果认为,我们这一物种通常本身消耗了至少是净初级生产力提供给所有陆上物种总能量的25%,而有的计算出来的结果竟达到了40%。保罗·埃利希概括了这些值得注意的数字背后的故事:“作为千千万万种物种之一,智人吸收了大约所有光合作用的产品的1/4供自己使用。”[3]
表6.1 从历史上看人均能量消耗(能量单位=1000卡路里/每天)
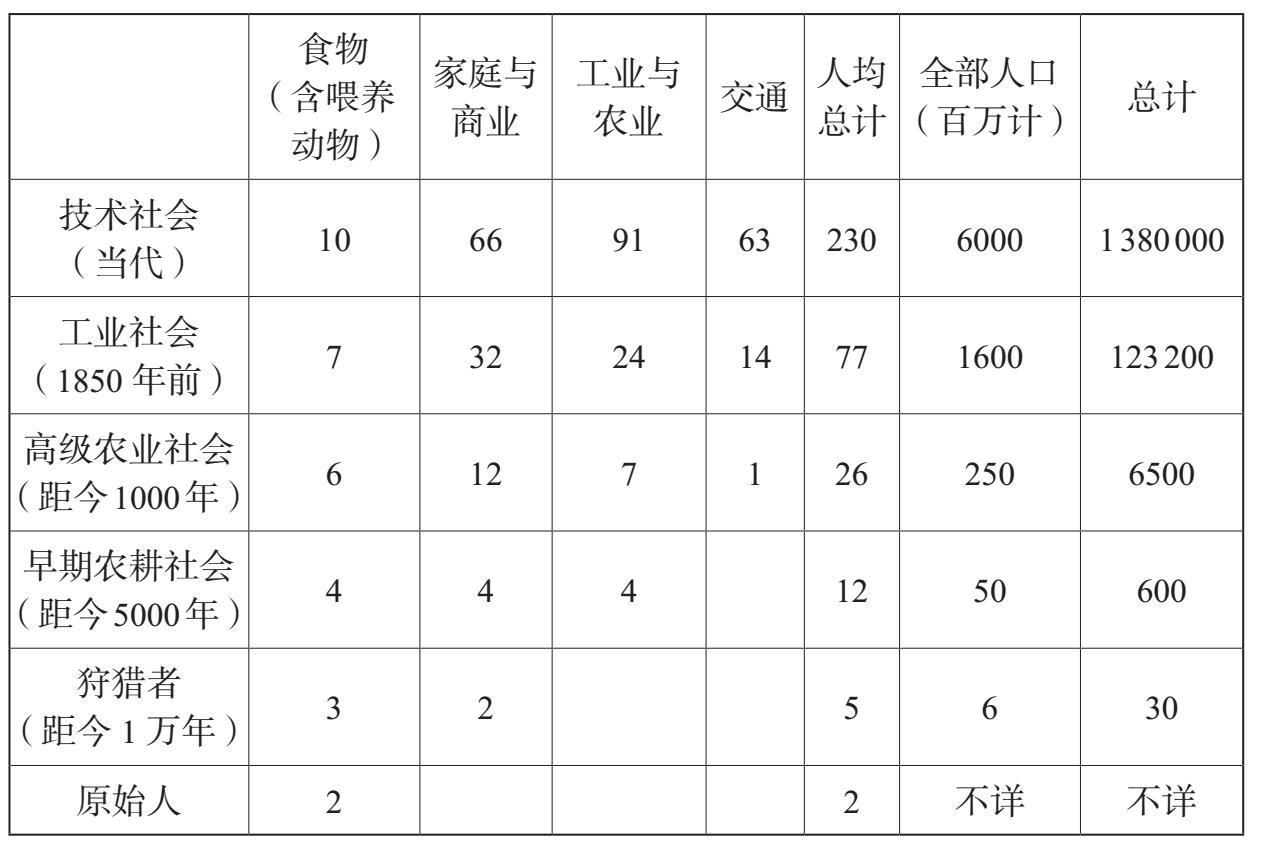
资料来源:I. G.西蒙斯(I. G. Simmons):《地球外貌的变化:文化、环境和历史》第2版(牛津:布莱克韦尔出版社,1996年),第27页
人类对能量的控制能力的逐渐增长,塑造着人类的历史以及许多其他物种的历史。它也使得人类能够以加倍的速度增长。表6.2、表6.3和图6.1总结了过去10万年里的人口增长。随着数量的增长,我们这一物种的生态范围也在扩张;人们发现,到1万年前,也可能早在3万年前,人类就居住在除了南极洲以外的所有大陆上。在旧石器时代,人类历史的特色主要表现为人类居住范围的不断扩大。在过去1万年里,不断增长的人类居住密度极大地塑造了人类社会的进化方式,人类学会在越来越大的共同体中生活,从农村到城镇、城市一直到国家。
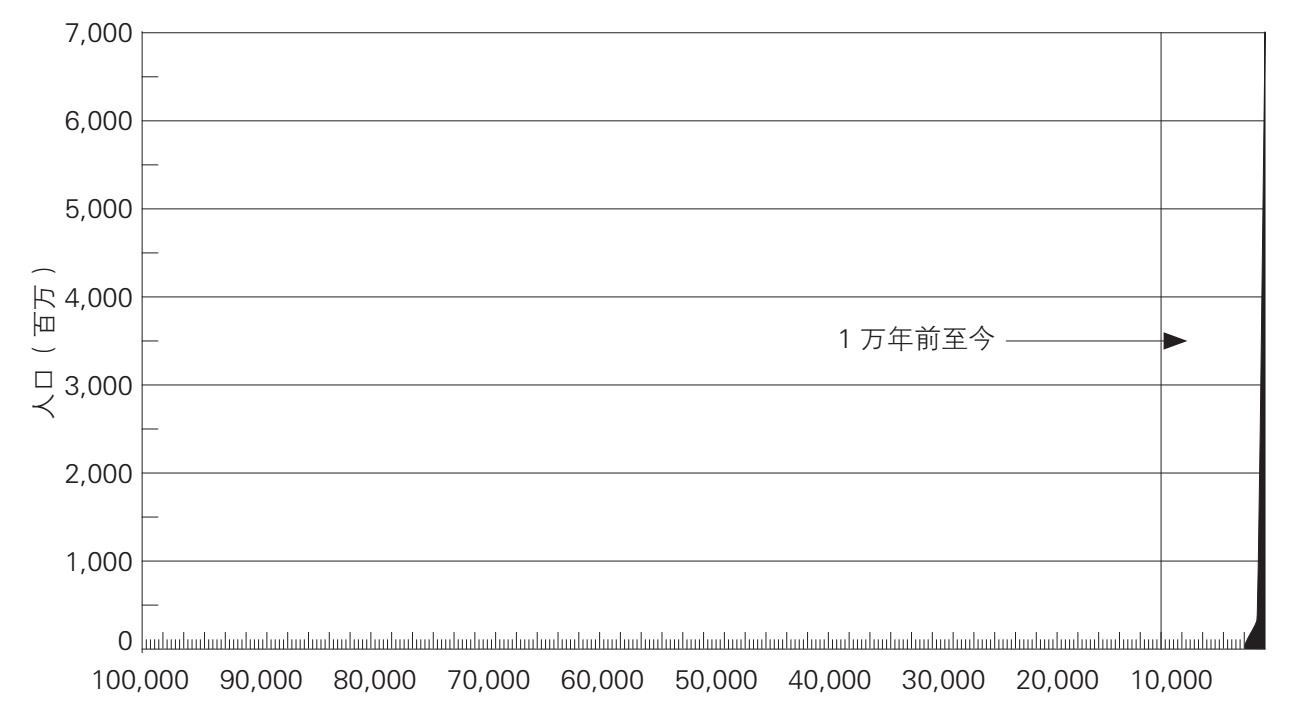
图6.1 人类的数量,10万年前(距今)至今(根据表6.2制)
表6.2 世界人口及增长速度,10万年前至今
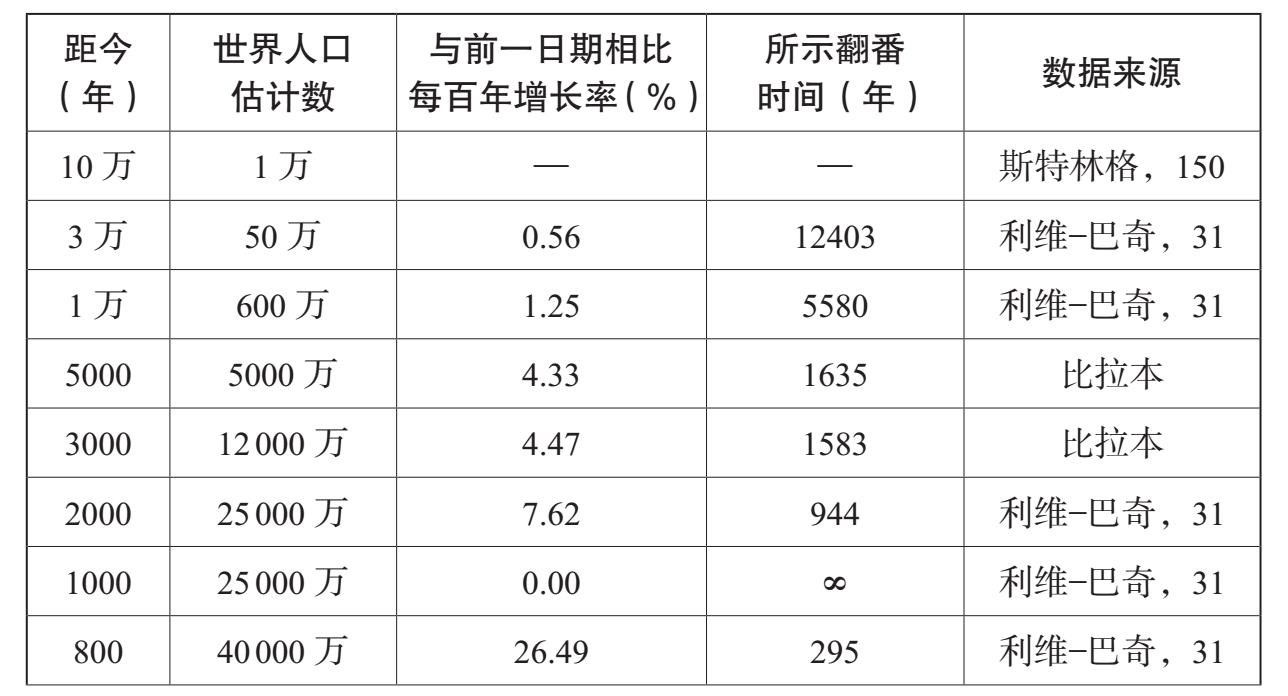
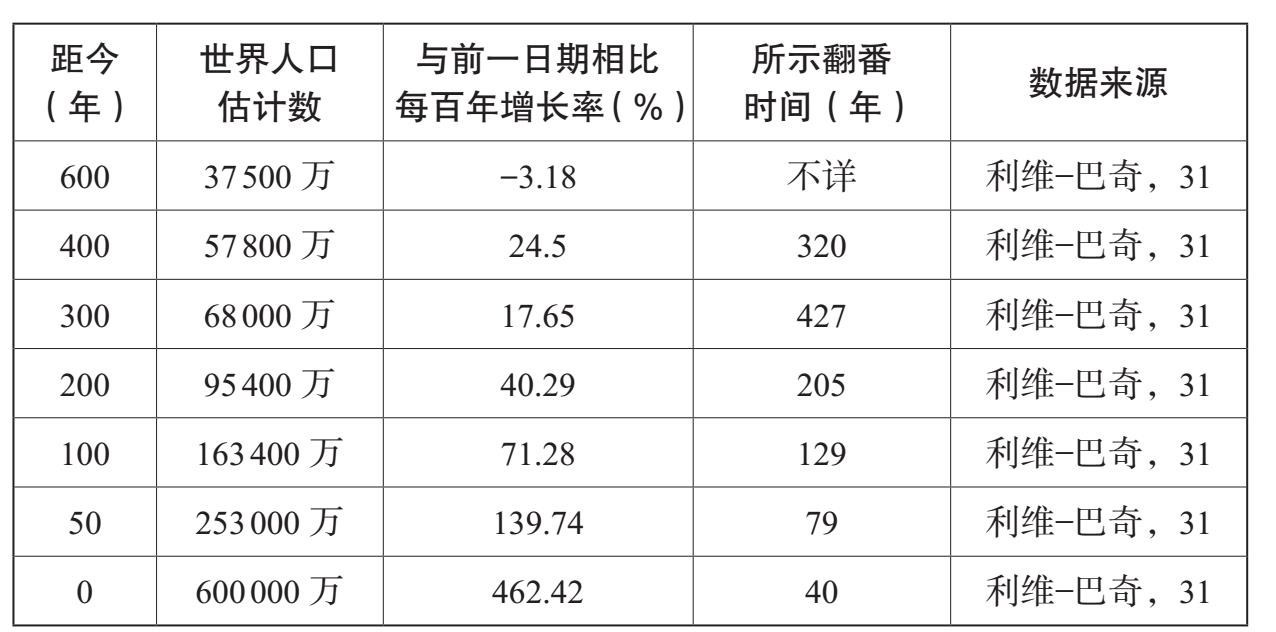
资料来源:比拉本(J. R. Biraben):《论人口数量的发展》,载《人口》第34号(1997年),第13—25页;马西莫·利维——巴奇(Massimo Livi-Bacci):《简明世界人口史》,卡尔·伊普森(Carl Ipsen)译(牛津:布莱克韦尔出版社,1996年);克里斯·斯特林格(Chris Stringer)和罗宾·麦凯(Robin Mckie):《走出非洲》(伦敦:凯普出版社,1996年)
表6.3 不同历史时期的人口增长速度
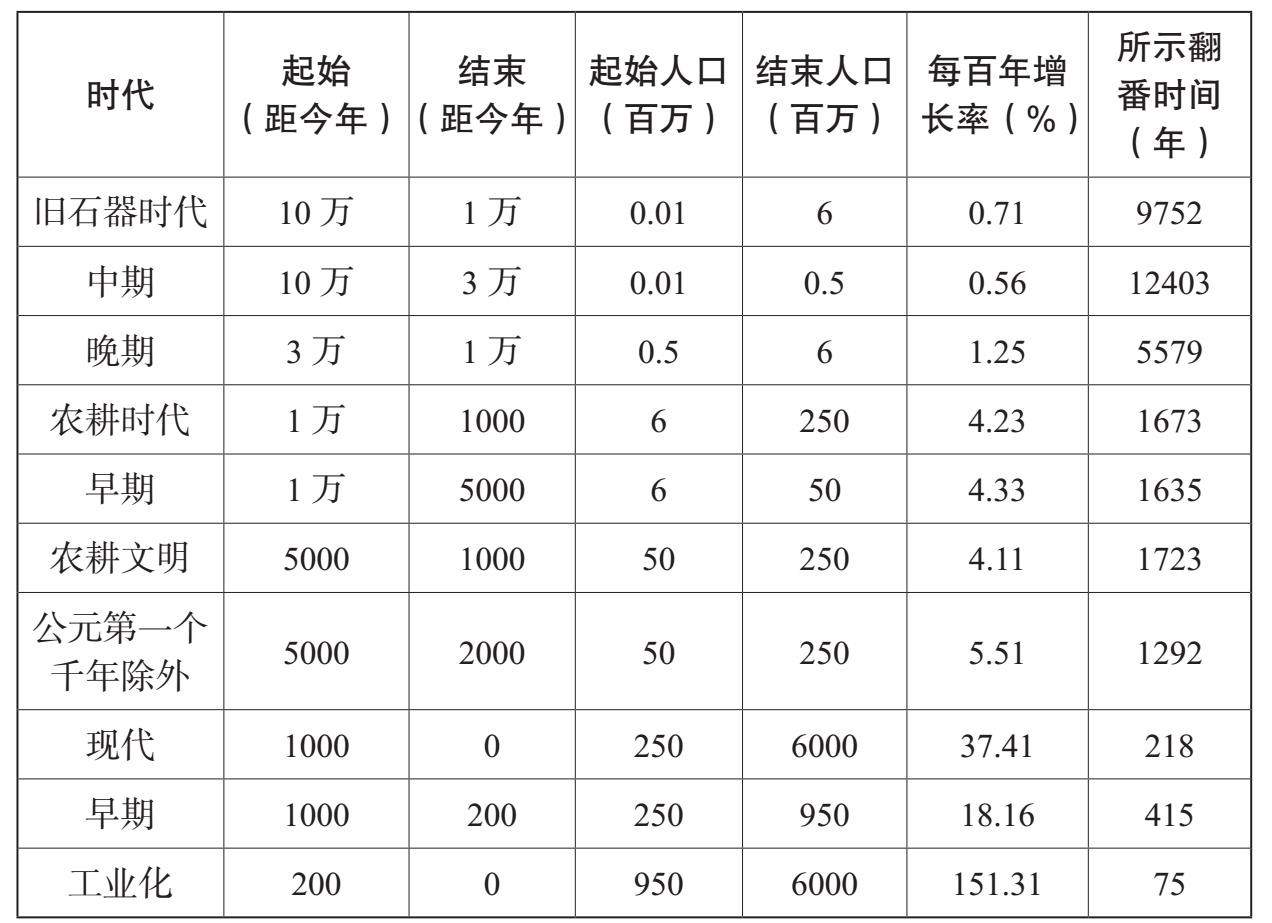
资料来源:表6.2
顾名思义,人类利用过的资源,其他物种就无法再利用。所以,当人类的数量上升时,其他物种的数量就会萎缩。驯化的动物,如羊和牛,以及从蟑螂到老鼠这些在无意中驯化的动物生养众多。但是更多的物种境遇都并不好,大量物种先后消失。这个过程同样是从旧石器时代开始的,那时人类的活动导致如尼安德特人那样的亲缘物种灭绝,也导致许多其他的大型物种灭绝,其中包括西伯利亚的猛犸、美洲的马和巨型树懒,以及澳大利亚的巨型袋熊和袋鼠。今天,人类活动所导致的物种灭绝速度正在加速。被认为正在受到“威胁”的物种有:4629种哺乳动物中的1096种(24%)、9627种鸟类动物中的1107种(11%)、6900种爬行动物中的253种(4%)、4522种两栖动物中的124种(3%)、25000种鱼类动物中的734种(3%),以及27万种高级植物中的25971种(10%)。[4]可以预想,由于物种灭绝速度加快,在不远的将来会有更多的物种消失。这些颇具说服力的数字可以衡量人类历史对这个星球的影响,因为古生物学家研究了过去超过6亿年的时间里的物种灭绝速度,而现在的灭绝速度似乎类似于那段时期中5—6次最激烈的物种灭绝的速度。[5]这就意味着人类历史的影响至少在今后10亿年中仍将可以看到。换言之,如果有外星古生物学家在这10亿年中造访这个星球并利用现代古生物学家的工具解读这个星球的历史,他们将会发现,我们这一物种的出现与其他物种的大灭绝,二者是同时发生的。
这些数字同样能够帮助我们衡量人类历史的独特性。再也没有其他的大型动物像人类那样繁殖、占据如此广阔的范围,或者说控制数量如此巨大的生态资源。(可能的例外是例如牛或兔之类作为人类生态队伍一部分而增殖的物种。)我们的历史甚至与我们最近的亲戚黑猩猩的历史也完全不同。尽管黑猩猩无论在遗传上、身体上、社交上和智力上都和我们非常相近,我们依然没有证据可以证明它们的数量、所占据的范围,或是它们的技术在过去的10万年里发生过巨大变化。事实上,这也正好是为什么人类能够说自己有“历史”,而认为黑猩猩也有历史的想法就显得有些奇怪了。许多动物物种并没有我们通常所说的历史,它们完成进化后,就会停留在最初的生态龛里,直到从化石记录上消失。整个物种的科或目,如恐龙或哺乳动物,能够说是有历史的,因为它们之中的不同物种能够以许多不同的方式进化,因而整个科的数量、范围和生态“技术”就发生了变化。但是对于单个物种来说通常并非如此。人类繁殖、行为的多样化,不仅是该物种,也是整个科或目所绝无仅有的——而且竟是在一个令人惊讶的短时期内实现的。显然,我们人类这个物种的出现,意味着某个重要门槛已被跨越。人类的历史标志着历史变化新规则的出现。所以,关注人类的历史并非只是我们在谱系上自我炫耀。人类这一物种的出现标志着地球历史上的一个重大转折点。如麦克迈克尔写道:“每个物种都是大自然的一个实验品。而只有这样一个实验品——人类——以此方式进化:他们逐渐积累的文化适应能力能够补充其生物上的适应性。通常为了在短期内取得某些收获(食物、领地和交配)的生物学驱动力,与通过不断增长的复杂的文化活动以满足这种内驱力的理智活动,二者前所未有地结合在一起,这正是人类这个‘实验品’与众不同之处。”[6]
关于人类出现的解释
若干年来,为了解释向人类的跃迁,人们提出了许多“最初的推动力”。从直立行走——由此解放我们灵巧的双手用来制造工具(达尔文理论的首选答案),到狩猎与食肉,到脑容量增大,再到人类的语言。此后的解释强调人类语言的重要性,认为其他因素只起到辅助的作用。
前文提到过一些稍微抽象的解释。所有的物种都是适应它们的环境的,但是大多数物种的锦囊妙计中,只有一二种适应环境的绝招。相反,人类似乎经常能够发展出新的生态绝招,找到从自身环境中汲取资源的新方法。用经济学家的术语来说,人类似乎有一种相当高的“创新”的发展能力。他们不是在达尔文理论的几十万年或是几百万年的时间范围内,而是在数千年到几十年甚至更短的时间范围内创新。我们面临的挑战是要解释人类是怎样、什么时候及为什么开始达到这种生态创造力的新水平的。如果我们能够解释这一被大大提高的能力,对我们解释人类历史的独特性将大有帮助。
我们已经看到,新形式的复杂事物的出现总是包括了大型结构的创造,在这个结构之中,那些原先独立的实体被联合在相互依存的新形式与相互合作的新规则之内。[7]根据这一提示,我们有望找到朝向人类历史的跃迁,它主要并不表现为作为个体的人类在自然界中的变化,而表现为个体与个体之间相互关联的方式的变化。这就意味着我们应该不仅关注基因、生理学或是早期人类大脑的变化,同样也应该关注我们的祖先相互影响、相互作用的方式的变化。
就像许多其他同种类型的跃迁一样,我们这一物种的出现也是相当突然的,从古生物学的时间范围上看,几乎就是一瞬间发生的事件。这就意味着我们有望找到独一无二的爆发点。在恒星的形成过程中,经过很长一段时间温度上升,直到氢原子开始结合的时候,这个爆发点就形成了。而人类的进化也是如此:经过数百万年进化而来的适应技巧,在跨过某道门槛的时候突然发生了某种转型。怎样来描述这道门槛呢?很明显它与学习能力的提高有关。许多动物,从扁形虫到蟾蜍,都在学习。但是大多数动物所学的大多数东西随着它们的死亡也就随之丧失了。当然,某些东西也会被传授下去。黑猩猩妈妈通过实际行动示范教会它们的孩子如何敲碎坚果或是摸索寻找白蚁,黑猩猩宝宝可能及时地再教给它们的孩子。但是我们知道还没有一种动物能够抽象地描述应该做什么——没有一个动物不用示范就能够解释怎样搜寻白蚁,或是不用走一遍就能描述一条小径,我们当然还知道没有一个动物能够描述诸如上帝、夸克或是幻觉等抽象的实体。过去与将来也是抽象的,只有现在是能够直接经历的。因此,没有符号语言的动物也就缺乏人类所具有的刻意去思考过去和想象未来的能力。这些都是严重的局限性。灵长目动物学家雪莉·斯特鲁姆(Shirley Strum)曾经多年观察过肯尼亚的一群狒狒,她称它们为“泵房帮”(Pumphouse Gang)。与其他群体相比,它们是狩猎艺术家,它们通常每天吃一次肉。但是,它们在一头特定雄狒狒的带领下,狩猎就格外成功。而一旦这头狒狒死去,它们就无法保存它的能力和知识。[8]
然而,人类的语言允许大脑之间更为精确和有效的知识传递。这就是说,人类能够更为精确地分享信息,创造生态和技术知识的资源共享池,也就是说,对于人类而言,合作带来的好处将逐渐超过竞争带来的好处。[约翰·密尔斯(John Mears)将人类称作“高度网络化的动物”。][9]此外,每个个体贡献给这个共享池的生态学知识在他或她死后能够长期保存下来。所以知识和技能能够不通过遗传而一代一代地积聚起来,每个个体能利用从许多前辈那儿积累起来的知识。因此人类的与众不同就在于他们能够集体学习。细胞式的思维(集中于个体的思维)很难看到这一点,但是在解释人类的独特性时,我们必须学会不要把黑猩猩个体和人类个体做比较(个体之间差异虽大,但并非不能改变),而是要和整个人群做比较。如果我们把人的大脑和黑猩猩的大脑做比较的话,我们是无法理解其间的差异的;只有当我们将黑猩猩个体的大脑与经过许多代、数百万人共同创造的大脑相比较时,我们才会开始领悟其间的差异。
集体知识的可能性改变了一切。麦克迈克尔写道:
累积的文化的出现是自然界一桩空前的事件。它产生了类似于复利的效果,允许连续的几代人在文化和技术发展的道路上不断前进。在这条道路上,人类大体上距离其生态根源越来越远。知识、思想和技术的传播代代相传,这给予了人类一种完全空前的能力,凭借这种能力,人类能够在完全陌生的环境中生存并且创造出他们所需和所想的新环境。[10]
集体知识赋予人类以历史,因为它意味着随着时间的流逝,人类可资利用的生态技能发生了变化。这个过程具有明确的指向性。随着时间流逝,集体知识的过程确保人类作为一个物种能够更好地从环境中汲取资源,不断提高的生态技能确保随着时间的流逝人口不断增长。对于集体知识的概述当然不能够预言这个过程的精确时间选择或是地理位置,也不能够预言它们可能发展到怎样的地步,更无法预知其确切的结果,但是这样的概述能够告诉我们在巨大的时间范围内人类历史的长期走向。
要感受集体知识的力量,我们只要做如下的想象就可以了:如果我们不得不从零开始学习一切事物,如果我们从家庭或共同体中接受到的东西只不过就是如何获得适宜的社会行为和饮食习惯,而这些多少只是年幼黑猩猩所得到的智力遗产而已,那么我们的生活将会是怎样一种情形。在我们的一生中能够发明我们身边多少种人造的事物(每一件都体现着积累的知识)呢?问这样一个问题便足以提醒我们,作为个体的生命在多大程度上依赖于许多代无数人的知识积累。作为个体的人类并不比黑猩猩或是尼安德特人聪明多少,但是作为一个物种,我们拥有巨大的创造性,因为我们的知识在一代人内部或是几代人之间都可以共享。总而言之,集体知识是一种如此有力的适应机制,以至于我们可以认为它在人类历史中所扮演的角色相当于自然选择在其他生物体历史中所扮演的角色。
为什么人类能够集体学习?这是因为人类语言的特殊性。人类的语言比非人类的交流方式更为“开放”。说它在语法上开放,是因为它在语法上的严格规则,使我们从极少数量的语言学要素,比如单词中,产生近乎无限多的含义。它在语义学上也是开放的——就是说,它能够传达范围极为广泛的意义——因为它不仅能够指涉我们面前的东西,而且能够指涉并不在场的实体,甚至是根本不会出现在我们面前的实体。我们能够运用符号,在我们的记忆中储存大量的信息块,然后我们可以用这些由符号组成的信息块构建更大的概念结构,符号使我们能够把具体的东西抽象化——就是说,从我们周边的事物中“蒸馏”出本质来。但是它们也能够指涉别的符号。所以它们能够浓缩和储存大量信息,就像我们称之为钱的符号筹码提供给我们一个存储和交换抽象价值的简洁而有效的方法。[11]符号语言让我们储存和分享那些千百万人累积起来的信息。总而言之,与前符号的交流方式相比,符号语言是一种有力得多的材料原动力。如泰伦斯·迪肯(Terrence Deacon)所指出的,前符号语言的交流方式“只能通过与所指涉之物存在的部分——整体关系来指涉某种事物,即使二者只是在习惯上相符合而已。尽管大量的对象和关系都可以用非符号表现方式,事实上,任何事物都可以用感官表现,但是非符号的表现方式不能表现抽象的或者触摸不到的东西”。[12]
如果这种论证是正确的话,那就意味着我们如果想要理解现代人类的进化,就必须先解释符号语言的出现。但是相当重要的一点是,我们必须明确地指出,在这一过程中没有什么事情是必然发生的。与恒星的形成有所不同,只要我们知道重力的作用以及核力的强弱,就能在统计学上预测它的变化,然而生物变化则比较随意,目标也不那么确定,正因为如此,活的生物体要比恒星更加变化多端。最后构成我们这一物种的元素不规则地随意地聚集到了一起,根本没有什么确定性,它们必须以这种特殊方式将自己聚集起来。最晚在10万年前,我们这一物种出现之后,人类的数量曾下降到少至1万个成年人,这就意味着当时人类就像今天山地猩猩一样几近灭绝。[13]这个统计不仅提醒我们进化过程的随意性,同时也提醒我们复杂实体的脆弱性。人类在地球上的出现完全是一桩极其偶然的事件。
证据和论证:构造人类进化的故事
创作一个关于人类进化连贯的、看似合理的故事是20世纪科学最伟大的成就之一。但是这个故事是怎样构成的?在更进一步研究人类进化的故事之前,我们必须检验一下用于组成这个故事的证据和论点的类型。
化石证据包括祖先物种的骨骼和它们留下的遗物:它们的工具、食物残余,以及它们在骨头或石头上所做的记号。现代古生物学者能够从一块骨头中收集到相当数量的信息。一块下颚骨不仅能够考证出一个物种,牙齿磨损程度能够告诉我们有关该动物通常所吃的食物,这些还能够告诉我们该动物所生活的环境以及它利用环境的方式。一块头骨能够告诉我们一个物种的智力水平。头骨的下部通常能够告诉我们该物种是两足行走还是四足行走:两足动物的脊骨从下面进入头骨,而四足动物的脊骨则是从后面进入头骨。脚趾骨显示了该动物是怎样行走的:如果大脚趾与其他脚趾是分离的(正如大多数的灵长目动物),我们就能确定它的脚仍旧是用来抓而不是专门用于走路的。通常我们只能找到为数不多的骨头。但是一具比较完整的骨骼,比如露西[唐·约翰森(Don Johanson)于20世纪70年代在埃塞俄比亚发现了她的40%的骨骼]则能够告诉我们更多。露西和她附近的遗物大约有300万年到350万年的历史,它们至少提供了那个时代早期人类的生理学上的详细证据。
人类活动的遗迹也同样重要。最为重要的是石器的发现,部分原因是因为不太耐用的材料——树皮、竹子等等制成的工具很少能够保存下来。用显微镜分析石器的切割边缘能够告诉我们它们切割的是什么;对石头来源的分析可以告诉我们它们的制造者是否主动从其他地方挑选出这些特殊的原材料;复原石器制作遗址中的石片可以告诉我们它们是如何制造的;工具制造的方法能够给予我们关于我们的祖先是如何思考的珍贵线索。对早期人类遗址中其他动物骨骼的分析能够告诉我们,我们的祖先是否吃肉以及他们如何猎获动物。例如,仔细分析骨骼上的切痕,有时会发现人类砍斫的痕迹覆盖在食肉动物的牙齿痕迹上。可以设想,这意味着早期人类是以被其他食肉动物杀死的动物为食的。利用不断发展的现代断代技术,所有这种类型的材料证据都能够或多或少地准确地断定其年代(关于放射性断代技术,参见附录一)。
但是化石记录是零散的。直到最近,我们还根本找不到一个关键时期中的化石,这段时期大约在400万年到700万年前,当时人亚科原人(通向我们物种的世系)从通向现代黑猩猩的世系分离出来。所以不得不用其他形式的证据来填补这个缺口。最近10年里最重要的证据之一是由分子断代法提供的。我们在第4章已经看到,很多进化的变化是随机的。这对于并不直接影响物种生存机会的那部分基因组而言更是如此,其中包括大量“垃圾脱氧核糖核酸”以及包含在所有人类细胞线粒体中的脱氧核糖核酸。这部分的基因组的基因变化是“中立的”——它不会影响已经发展了的生物体。垃圾脱氧核糖核酸中的变化因而就像是在洗一大副纸牌。幸运的是,这种随机过程服从于普遍的统计学规律。如果你取一副按顺序排列的新牌,洗若干次牌,统计学家就能够根据这副牌与最初状况之不同的程度到底有多大,从而粗略地估算出洗了多少次牌。牌的数量和洗牌的次数越多,这种估算就越精确可靠。
在一篇最初发表于1967年的文章中,两位在美国工作的生物化学家文森特·萨里奇(Vincent Sarich)和艾伦·威尔逊(Alan Wilson)论证,很多遗传变化皆遵从类似的规则。[14]因此,如果我们取两个现代物种,计算出它们脱氧核糖核酸序列的差异,我们就能很准确地估算出这两个物种是何时从共同的祖先那里分离出来的。这样,脱氧核糖核酸的进化能够提供某种遗传时钟。这个观点最初受到了嘲笑,部分是因为很多人认为进化就是适应环境,这是自然选择的基本原则,它意味着进化的发生并不是按照在统计学上可预见的方式进行的。尽管如此,如今人们普遍同意,许多变化其实确实是随机的——无论如何,这种断代法所得出的结论与其他类型的证据是非常吻合的。这种基因比较方法现在通常用于理解不同物种之间的关系,尽管仍然存在一些问题。例如,如果它被当作时钟一般精确的话,很明显的是,不是所有的基因变化的发生都伴随着有规则的必然性。但是这些方法在许多方面都是极其宝贵的,特别是在人类进化的研究中。[15]
萨里奇和威尔逊首先告诉我们的是,一般而言,我们与黑猩猩之间的关系要比我们曾经想象的更近。在20世纪70年代,人们普遍认为人和猿的两个发展序列至少在1500万年前,甚至可能3000万年前就分离开来了——对于那些并不乐意承认我们与黑猩猩之间亲密的血族关系的人而言,这是一个比较合适的距离。然而,现代人类的脱氧核糖核酸与我们这个现存最近亲属的脱氧核糖核酸的差异只有大约1.6%。这就是说,我们的脱氧核糖核酸有98.4%是与现代黑猩猩一样的。这意味着我们的历史和黑猩猩的历史的所有变异必须根据在我们遗传材料与黑猩猩遗传材料之间的1.6%的差异而做出解释。对哺乳动物遗传变化的速度可以进行比较,这是因为我们知道,大约6500万年前当恐龙趋于灭绝的时候,哺乳动物各物种之间开始迅速分离。但是人类和黑猩猩之间的差别,仅相当于哺乳动物各主要物种之间的差别的10%左右,这意味着人类和黑猩猩是在大约500万年到700万年前相揖别的。这意味着在它们分离之际,有一种动物是现代人类和现代黑猩猩的共同祖先,尽管它有可能看上去与这些至今仍存在的物种有些不同。这段时期里化石记录的缺乏意味着我们对这位祖先几乎一无所知。[16]但是我们能够确定,这样一种动物确实存在过——否则我们就不会存在!类似的观点认为人类和大猩猩在800万年到1000万年前有着共同的祖先,而人类和猩猩则在1300万年到1600万年前有着共同的祖先。
根据对气候变化和现有动植物的分析,我们也能够知道许多关于我们人亚科原人祖先进化的环境。在过去几百万年里,全球气候为冰川时代不稳定和不可预知的气候变化所主宰(参见第5章)。这些变化改变了动植物的生活环境和外界环境,有利于具有高度适应性和能够利用各类生态龛的更为多样的物种。多面手或者是“长得快的”物种能够更好地适应生态破坏,譬如现代人类,他们可能就是冰川时期的典型产物。[17]
多方面的技术联合起来使我们能够描述人亚科原人的进化以及他们的生存环境,但是要描述他们的行为则要棘手得多。化石能够告诉我们有关生活方式的一些事情,但是为了得到更进一步的了解,我们不得不依赖与其他可能有近似生存方式的物种进行现代的类比方法。从珍·古道尔(Jane Goodall)和黛安·福西(Dian Fossey)开始,最近10年来的研究人员研究了野生类人猿(Great Apes)的生活,关于它们如何生活以及它们的社会关系、性关系和政治关系,[18]现在我们已经有了很多了解。这样的研究能够提示我们早期人类可能是如何生活的,但也有可能会误导我们,因为不同类型的类人猿,甚至同一类型的类人猿的不同群体,也会过着一种完全不同的生活。例如我们最熟悉的普通黑猩猩(Pan troglodytes),生活在相互关系非常密切的雄性所统治的共同体中,而其他群体的雌性便加入到这个共同体里面来。雄性黑猩猩构成不同等级,但是这些等级是可以改变的,雌性能够与数个雄性结合,在这种情况下,这些共同体的性关系和政治生活变得相当复杂。大猩猩则恰恰相反,它们通常生活在较小的群体里面,这些群体通常由数位雌性和一或两位雄性所组成。猩猩是最为孤独的,它们只有在配对时才会在一起。所以要确定灵长目社会中具有哪些相似性,从而使我们了解早期人亚科原人的社会,这并不是件容易的事。
同样,其他的类比方法也曾极大影响到人亚科原人进化的研究:与现代“食物采集”社会相类比。[19]人类学者经常提醒古生物学者,现代食物采集社会是非常现代的——无不以某种方式受到了现代社会的影响。所以,将人亚科原人社会或是早期人类社会的学说建立在这些类比基础上,可能是一种冒险。然而,由于现代植物采集社会的技术和社会结构当然要比现代城市共同体的技术和社会结构更加接近于早期人类,人类学家的警告往往被忽视了。现代的研究,如对非洲南部的桑人(San People)的研究有助于我们构筑一些似乎合理的模式:早期人亚科动物或人类如何狩猎、男女之间的相互联系怎样、他们会玩哪种类型的角力游戏。可能最重要的是,他们提醒我们,相对于现代城市居住者来说看似简单的社会也有其复杂而高度发展的一面。毕竟,在非洲南部或澳大利亚的沙漠,或者是西伯利亚的冻土地带,使用石器时代的工具成功地生活长达数千年之久并非易事。
最后,如今对其他物种如何进化的理解也被用来构建人类如何进化的模式。举例来说,经常可以发现,一个新的物种与它所进化来的物种中不成熟的个体极为相似。这个过程被称作幼态持续。它是通过一种物种中控制生命循环的基因开关的微小改变产生的。这样的变化能够产生第二种和第三种连续性效应,从而引起重大的进化。有人提出,在许多方面,人类较之成年黑猩猩更像幼年的黑猩猩;这种相似性暗示了我们可能是通过某种形式的幼态持续而进化的,而现代黑猩猩则可能保留更多我们成年共同祖先的成分。同样,现代进化论的研究证明,进化的发生通常是一阵阵的。如果由于环境的变化而出现一个新的生态龛,它经常会迅速被大量十分相似的物种所填补(“迅速”在进化过程中往往意味着几十万年甚至几百万年),它们之中大多数可能逐渐灭绝了,只留下一条或两条存活的序列。这个过程被称为适应辐射,大体而言,每个辐射似乎都与某种特殊的生态计谋有关。我们将会看到,在我们的祖先物种中似乎也有若干个适应辐射,我们现在可以认识到,其中每个适应辐射都在最后形成我们人类的进化包上增加了一些新东西。[20]
所有这些类型的证据都被用来构建这个人类如何进化的现代论述。这个论述还远非完美无缺,但是与10年前的论述相比,它更加丰富,建立在更多证据的基础之上。
灵长目动物和人科的辐射
我已经论证了符号语言的进化标志着通往人类历史的重要门槛。但是,我们的祖先如果不曾拥有能使他们利用符号语言所赐予优势的其他特质的话,符号语言也无法发挥这么大的作用。在所有这些预适应性之中最重要的是社会性、预先存在的语言技能、直立行走和灵巧的双手、食肉与狩猎、长时间的儿童学习期,以及大容量的前脑。在这里,我们将回溯这些偶然的过程,在这些过程中,各种因素进化并且结合成为一个构成我们这个物种的系列特征。
灵长目动物的遗产
我们具备许多以其他灵长目动物命名的特征。[21]大多数灵长目动物曾经栖居树上。在树上生活的动物必须视觉良好,否则它们就会掉下来。所以,所有的灵长目动物都具有良好的立体视觉。味觉对灵长目动物来说是次要的,不如味觉对于狗那么重要,这也是为什么许多灵长目动物都有小小的嘴和平平的脸。在复杂的、三维的环境里的视觉信息需要经过许多处理,因此大多数灵长目动物拥有相对于它们的身体大小而言相当大的大脑,而且整个灵长目动物都具有一个特征,就是大脑的容量不断增长。较大的大脑通常表明较长的寿命——也许是因为这表明它们更加依赖于学习,而(原则上)年龄越大,学到的越多。在树上生活还需要四肢灵敏,所以大多数灵长目动物都有能够很好地抓握和控制物体的四肢。实际上,这意味着它们的大拇指和大脚趾能够与其他手指和脚趾相对。与居住在陆地上的物种相比,在树上生活还促进了前后肢的分工。尽管许多灵长目动物用前肢或用后肢都能抓住东西,但是它们的后肢趋向于专门用于运动,而前肢则专门用于抓握物体。
人类属于旧大陆(Old World)灵长目动物里特殊的一群,被称为人猿总科。包括人和猿——黑猩猩、大猩猩、猩猩和长臂猿——还有它们现在已经灭绝了的祖先。分在这一总科的生物体的最古老化石只超过2000万年,这就意味着它们出现在地质学家所称的中新世早期(大约距今2300万——520万年前)。这些遗存属于一种叫作原康修尔猿的物种。[22]尽管类人动物可能在非洲进化,但是早至1800万年前类人动物的遗存同样也在从法国到印度尼西亚的欧亚大陆南部发现。类人动物有很多不同的群体,在一段时间里,它们可能比旧大陆里其他猴类的数量还多。它们的迁移为适应辐射提供了一个典型事例。
化石记录提供的解释并不足以说明,进化技巧就是定义类人动物的最佳方式,尽管不断增长的体型、越来越灵巧的手工操作能力、越来越大的大脑,以及愿意离开树冠等可以算作这些技巧的一部分。所有这些都是我们与现存灵长目总科里的其他成员所共有的特征。
两足直立行走和最早的人亚科原人
人亚科(Homininae)是人科动物(Hominidae)大猿的一个亚科。人亚科原人只包括我们自己的直接祖先。他们的历史开始于600万年到500万年前从中新世到上新世的过渡时期。这个历史开始于基于分子测定年代技术的认识,即大约600万年前,在非洲的某地存在一种动物,它是现代黑猩猩和现代人类的共同祖先。从那以后,经过一系列的适应辐射,大量不同种类的人亚科原人出现了——可能多至20或30种。30年前的困难在于找不到任何人亚科动物的遗存,而今天的难点是确定我们目前所知的这些物种中的哪一支沿着这条序列进化到了现代人类。
对一个现代古生物工作者来说,最神圣的任务是要发现黑猩猩和人类的这一共同祖先的遗存。很可能这个物种,或者说与它相近的某个物种已经被发现了。2000年,由法国和肯尼亚的考古工作者组成的一支队伍,在内罗毕北部发现了大约600万年前的一种生物遗存,新闻界迅速称其为“千禧年人”。[23]但是它的真实身份仍然不能确定。它的外貌非常像猿,以至于许多古生物学家在需要区别黑猩猩和人亚科原人之际,更倾向于把它放在黑猩猩一边,而不是人亚科原人一边。对于另一个可能最古老的人亚科原人,卡达巴地猿始祖种(Ardipithecus ramidus kadabba),人们也曾提出过类似的批评。2001年7月的《自然》杂志报道,一队美国考古学家在埃塞俄比亚的东非大裂谷发现了该物种的遗存。[24]这些遗存的年代在580万年到520万年前之间。它们包括一块脚趾骨,它的形状表明这一生物是两足直立行走。目前,大多数古生物学家都同意,人亚科原人同猿的决定性区别在于直立行走:所有已知人亚科原人都是直立行走的,而所有已知的猿都不会直立行走(尽管黑猩猩能够站立很短一段时间)。[25]所以断定这些早期标本是否真正两足直立行走将是非常关键的。就目前而言,证据是有歧义的。
对这些发现的价值的讨论变得错综复杂,因为事实是尽管有许多学说,但是谁也不能确定为什么这一物种会进化到两足直立行走。[26]一些人将焦点放在气候变化所起的作用。2000万年前,非洲大陆相对较平坦,其赤道地区甚至还完全为热带雨林所覆盖。但是大概从1500万年前开始,非洲大陆构造板块开始分裂为两半,沿着东非大裂谷的构造活动创造了一连串的高地,峡谷沿着大陆的东部向北部和南部运动。裂谷劈开了地壳,为化石的寻猎者提供了一个愉快的场所。但是能够解释这里存在着人类化石的乃是山脉,因为它们严重影响了东部大陆的降雨,使得这片区域比西部的更加干燥。伊夫·科庞曾经提出,这种干旱将一些物种赶入较少为森林所覆盖的地形中去,在那里,它们不得不在树木之间移动较远的距离来寻找它们所习惯的那类食物。这可能鼓励了直立行走姿态的进化,因为黑猩猩指关节行走方式的特性并不适合于长距离跋涉。而对这个看似有希望的理论来说非常不幸的是,最近发现的一些早期人类化石,包括那些卡达巴地猿始祖种的化石都是在几乎为森林所覆盖的环境中被发现的。[27]
两足直立行走可能使得人亚科原人在空旷地区看到更远处潜在的食肉动物。与黑猩猩特有的用指关节行走相比,也可能能效更高,使得早期人亚科原人能够在更大的区域内搜寻食物。在没有遮阴的环境下直立行走也可能减少了皮肤直接暴露于阳光的面积,从而降低了正午烈日的伤害。在各式各样的压力面前,喜欢直立行走的个体具有更强的优势。(最后一条论据也能够解释为什么人亚科原人在进化到某一点的时候,就变得不像其他大猿那么多毛了。)与黑猩猩进行比较是有启发性的,正如科庞指出的,黑猩猩在三种情形下试图站立:“看得更远,保护自己或是发动进攻——因为站立起来就可以腾出双手,抛掷石块——还可以为后代携带食物。”[28]
无论是什么原因导致直立行走,化石证据——虽然相当罕见——却依然表明,在200万年内,有一些两足行走的物种出现了。其中包括卡达巴地猿始祖种——1994年在埃塞俄比亚发现了它的遗存——在内的物种,其年代大约为440万年前。这些早期人亚科原人物种,不管它们是什么,构成了人亚科原人历史上最初的主要适应辐射,而它们的成功可能和两足直立行走的优势密切相关。
南方古猿
另外两种人亚科原人辐射与古生物学家所指的南方古猿属的种联系在一起。
所有南方古猿都是两足直立行走的。我们是根据骨盆的构造、手臂和腿部的相对长度和脊骨进入颅骨的切入点(从下面而不是从后面)而知道这一点的。现在已知最古老的南方古猿是南方古猿湖畔种,1995年其遗存发现于在肯尼亚北部的图尔卡纳湖地区(Lake Turkana)。它们的年代测定为距今420万年前。[29]最著名的南方古猿的残片是美国古生物学家唐·约翰逊(Don Johanson)于20世纪70年代在埃塞俄比亚发现的。他找到了一具两足直立行走女性40%的骨架,将她命名为露西(Lucy)(据报道是以甲壳虫乐队的流行歌曲《钻石天空中的露西》命名的)。露西大约有1.1米高,尽管附近的其他遗存最高可达1.5米。所有这些遗存都距今370万年到300万年,它们通常被归类为南方古猿阿法种,以它们所被发现的埃塞俄比亚阿法地区(Afar valley)命名。[30]1998年,在南非发现了更为完整的南方古猿的骨骼,并且还有头骨。其年代在距今350万年到250万年间。玛丽·利基(Mary Leakey)发现的著名的拉托利(Laetoli)脚印更为古老,其年代至少在距今370万年到350万年前。这些脚印是由三个南方古猿留下的,其中两个是紧挨着并排走的,第三个是带队的。他们显然是在通过热火山灰时手搀手走过的。这些令人吃惊的脚印直接证实了其他化石遗存所间接提示的东西:我们所知的最古老的人亚科原人是两足直立行走的。1995年,在东非大裂谷西边乍得工作的考古工作者发现了一种新的物种的遗存,南方古猿羚羊河种(Australopithecus bahrelghazali),生活在距今300万到350万年前。很明显,在东非大裂谷的两边都曾经生活着南方古猿。20世纪所发现的数百个南方古猿个体的遗存遍布从埃塞俄比亚到乍得再到南非的很大一块区域。

图6.2 复原的露西
露西是大约320万年前生活在今埃塞俄比亚哈达(Hadar)河谷的南方古猿。她身高约1.1米,脑量与现代黑猩猩相同。选自戈兰·布伦哈特(G. Burenhult)5卷本《图说人类历史》第1卷:《最早的人类》(旧金山:哈珀旧金山出版社,1993年,第1版)。维尔登·欧文(Wenldon Owen)私营公司/布拉伯克(Bra Bocker)版权所有(1993年)。哈珀·柯林斯出版社惠允复制
尽管南方古猿已经两足直立行走,但是解剖学特别是对它们的手的详细研究显示,它们非常适应于栖树生活,它们依然不能像现代人类那样高效行走。更重要的是,它们的脑容量较小,在380—450毫升之间。这和现代黑猩猩300—400毫升的脑容量以及现代人类平均1350毫升的脑容量形成了鲜明对照。人亚科原人发展序列的第一个显著特征不是聪明的头脑而是直立行走(参见图6.2)。
我们有足够的理由认为,我们的世系可以追溯到早期形式的南方古猿。但是同样以一种非常独特的辐射方式出现了另一种群的南方古猿,用古生物学的语言来说,比阿法种南方古猿看上去要“粗壮”得多。它们存在于距今300万到100万年前之间,有时被划分为一个独立种类:傍人(Paranthropus)。它们与众不同和标志其与我们不同的进化序列的地方,就在于它们特别进化出了强壮的颚,能够研磨坚硬的、纤维性的植物食物。因此它们的头骨粗壮,带有夸张的嵴,能够为强有力的咀嚼肌肉提供支撑点。
对于南方古猿的生活方式我们能说些什么呢?如果从它们通常所吃的食物说起,似乎大多数南方古猿主要依靠的是其祖先在森林环境中所吃的那些食物。它们的牙齿适应了研磨坚硬的或纤维性的水果的外壳、树叶和其他植物。然而,它们可能偶尔也吃肉,因为直接的观察表明,现存大多数灵长目动物经常食肉,而且尽可能多地食肉。[31]它们有时猎取体型较小、较弱的动物(包括其他灵长目动物),有时食用自然死亡或是被其他食肉动物杀死的动物的腐肉。但是大体上,南方古猿是素食动物。
与占据类似生态龛的现代灵长目动物的类比分析表明,南方古猿可能是以小家庭团体的方式生活的,它们集体行走,分别为自己采集食物。没有证据表明它们的语言能力比现代黑猩猩更强。这并不意味着在它们的群体中就没有政治或是交流了。在许多现代灵长目动物的社会中,雌性和雄性可能形成统治阶层并花费大量时间来处理——也可能是思考——集团政治。就像现代的黑猩猩,南方古猿可能通过手势、声音和梳理毛发等活动来进行交流。但是黑猩猩和南方古猿都没有必需的发声器官或是智力,不能精确地传达抽象信息。
对于和现代人类关系相近的灵长目动物社会的研究,则为最早的人亚科原人社会的性质提出了相反的观点。在遗传学上,我们和黑猩猩最为接近,而其中最著名的成员普通黑猩猩(Pan troglodytes),以关系相近的雄性黑猩猩为纽带过着群体生活。雄性与它们出生时的群体在一起生活,而雌性则离开它们出生时的群体。但是大多数南方古猿的种并不像黑猩猩,它们似乎是两性异形的(也就是说,雄性要比雌性大得多)。这就意味着南方古猿“社会”在某些方面可能更加接近大猩猩。[32]在大猩猩那里,雄性体型较大是因为它们需要相互竞争以便占有雌性,这就确保了最大的雄性能够最多地繁殖后代。其结果就是组成一个社会,其中有一个占统治地位的雄性,或许还有另外一个年轻雄性,与若干雌性以及它们的孩子组成最多可达大约20个个体的群体一起行动。或许我们应该预想到一个多少介于这些结构之间的世界。它可能是一个比现代黑猩猩群体稍小的世界,关系相近的雄性为统治地位和占有雌性而相互竞争。居于统治地位的雄性也许可以占有数位雌性,但是这种权力并不是独享的。南方古猿也许就是生活在这样一个世界里:雄性为吸引雌性伙伴多少要经常相互竞争。不过,在这个虽然竞争激烈然而关系相近的雄性世界的生殖核心中,如同现在许多灵长目动物一样,仍然存在着由母亲和它们的孩子所组成的更小的但是更近的单元。我们知道,黑猩猩妈妈对自己的孩子有着持久且明显的亲情关系,而雄性则对抚育孩子的工作以及父子关系毫无兴趣。总而言之,几乎没有什么能够表明,南方古猿在生理学或是在生活方式上与今天的猿类有什么根本不同。
使用工具和食肉:能人
对于古人类专家而言,在坦桑尼亚北部的塞伦盖蒂平原(Serengeti Plain)上绵延50千米宽的奥杜韦峡谷(Olduvai Gorge),作为东非大裂谷的一部分,乃是一个特殊的地方:这里的发现为我们人类的进化起源于非洲提供了最好的证据。在这里,1960年乔纳森·利基(Jonathan Leakey)——现代人类进化研究的先驱者之一路易斯·利基(Louis Leakey)的儿子——发现了一具大约1.4米高的人类遗骸。路易斯·利基称它属于人类同属(人属),因此命名它为“能人”,或者叫“巧人”(Homo habilis)。这使它成为包括现代人在内的人属中最古老的一个成员。
尽管许多人类学家觉得这具遗骸只是属于南方古猿中一种特别纤弱的类型,但有两个因素使利基相信该种更应该是“人”。首先,他发现了能人系统制造和使用石器最早的证据。值得注意的是,在这些活动中所包含的技能显然要比早期人亚科原人的更复杂。其次,能人的脑容量要比南方古猿大得多,在600—800毫升之间。能人似乎是一个会使用工具、学习的动物,就像现代人类一样,因此在大约230万年前这一新种的出现可能标志着人类历史的真正开始。现代人类学家保留了利基的命名法,毫无疑问能人显示出与众不同的特征,其中一些可能是由于始于250万年前的气候变冷变干所导致的生态变化所引起的。例如,能人制造的石器显示了“惯用手”的迹象,暗示大脑有了左右分工;这也可能是提高语言技能的一个必要前提。[33]然而,近期对越来越多的能人的遗骸和遗址的研究表明,能人与现代人类之间在智力能力和生活方式上的鸿沟要比利基所想象的大得多。[34]
对于能人态度发生转变的部分原因是因为对于能人使用工具的迹象,现代的古生物学者没有像利基那样留下深刻的印象。我们现在知道,许多动物都使用某种工具,除了人类之外,黑猩猩比其他动物更会使用工具。例如,我们已经发现黑猩猩会把棍子插入白蚁堆中,然后迅速抽出棍子就能吃到仍然爬在棍子上的白蚁,还有一些黑猩猩甚至会用石头砸开坚果。然而,能人似乎用一种更需要计划和远见的新方法来使用工具。古生物学者把他们的石器以发现大量此类石器的奥杜韦峡谷命名,称之为奥杜韦文化(Oldowan)。这些石器有着十分特殊的形式,在考古学上的记录延续了近200万年,几乎一直到25万年前(参见图6.3)。它们主要由大石块组成,通常是坚硬的玄武岩或是石英岩形成的河里的鹅卵石,用石“锤”打制成小的碎片产生一或两个切割边缘。
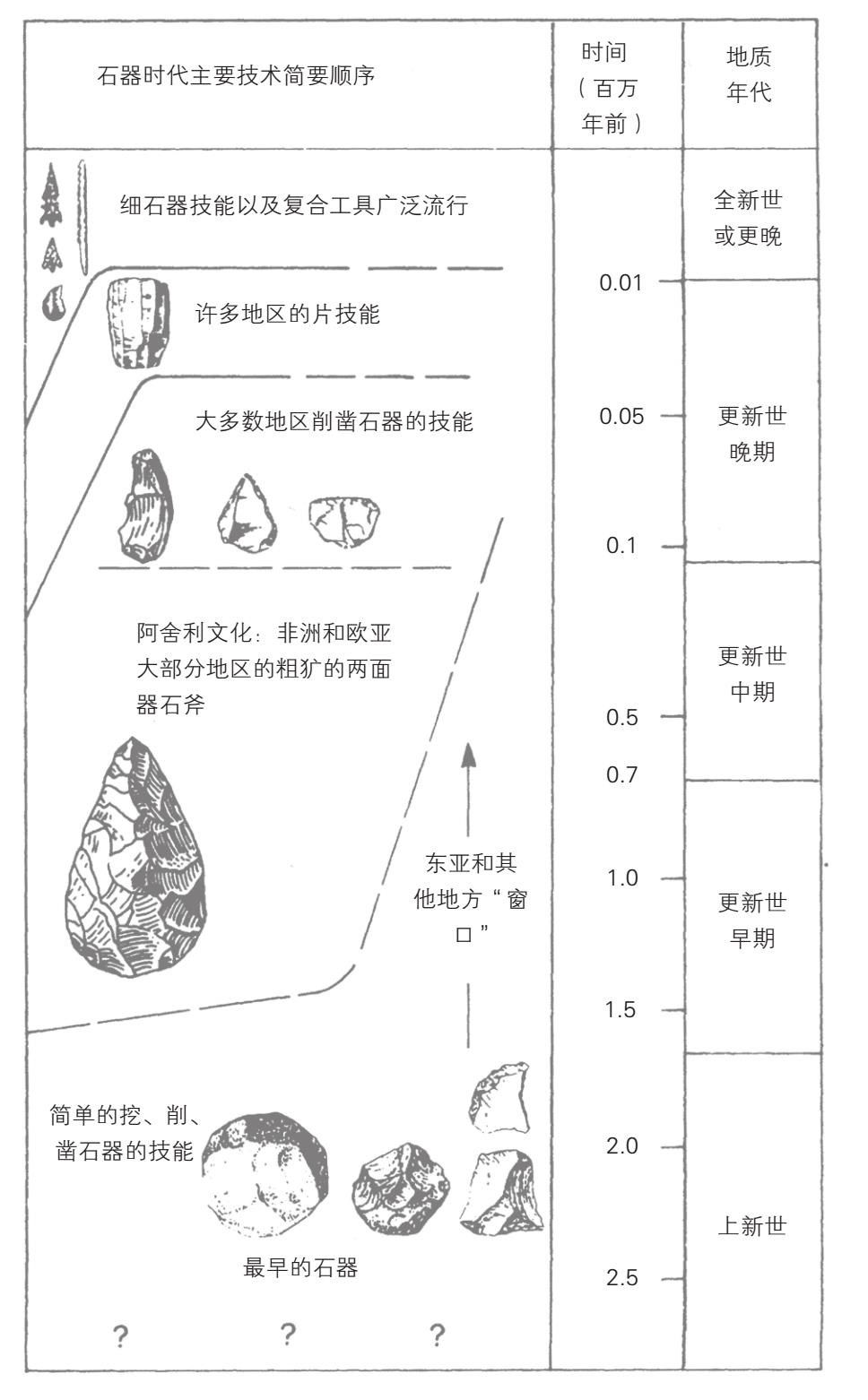
图6.3 250万年来石器的进化
选自史蒂文·琼斯(Steven Jones)、罗伯特·马丁(Robert Martin)和戴维·皮尔比姆(David Pilbeam)等编:《剑桥百科全书·人类进化》(剑桥:剑桥大学出版社,1992年),第357页。剑桥大学出版社授权复制
制造此类工具需要比黑猩猩制造简单工具更多的计划和经验。现代敲碎石块的试验表明,原始石块需要精心挑选,精确打制。事实上,制造石器所需要的精确技能乃是额前部皮层所特有的,这一部分大脑在人类进化过程中最为显著地发展了。使用工具的进化很可能经过一个被称为鲍德温适应过程(Baldwinian adaptation,以19世纪美国一位心理学家的名字命名,他首先系统地描述了这个过程)。这种进化形式似乎将达尔文学说和文化因素结合起来,因为行为变化导致一种动物生活方式的变化,由此产生一种新的选择性的压力,随着时间的流逝,便导致了遗传上的变化。例如,学会了新的行为而能够在寒冷环境中居住的动物,可能最终在遗传上进化毛皮适应它们的新环境(如猛犸或双角犀牛)。在人类中,能够放牧驯化动物的群体经过许多代人最终获得了很高的消化牛奶的能力,因为普遍发生了罕见的突变,延长了成年人消化牛奶的乳糖酶的产生。可能以同样的方式,那些最善于制造和使用工具的人亚科原人获得了自然选择的优势而比其他人拥有更多的后代,因此他们智力上的技能很快合并到整个物种的遗传结构中去了。如果是这样的话,那么工具的使用便是大脑成长的起因,也是其结果,这是一个正反馈过程。
那么石器是用来派什么用处的呢?现代的试验显示了奥杜韦文化期的砍砸器能够成功地打碎骨头或是对木头进行粗加工。但是打制时剥落下来的碎片可能比其中心部分更为重要,因为这些碎片形成小而锋利的薄片,可以用来屠宰和切割。所以我们能够想象能人个体和群体在采集植物时随身携带着小砾石,需要时就从这些砾石上打制一些薄片来用。用显微镜观察这些石器的边缘,我们可以看到奥杜韦文化的石器有多种用途。可能其最重要的作用是用来获得更丰富多样的食物。它们能够用来挖掘植物块茎,舍此别无他途。更重要的是,他们制作的砍砸器和薄片能够用来吓跑来掠夺动物死尸的其他食肉动物,获得大型动物的骨髓,以及切割动物尸体。从能人的牙齿来看,不管他们是怎样获取肉食的,他们吃的肉食肯定比南方古猿更多。丰富的食物原料可能提供了一些支持更大脑量所需的新陈代谢的能量,如果食肉果然能使肠子缩短,因而减少加工和消化食物所需要的能量,那么情况就更是如此了。食肉可能也导致更复杂的社会生活,最近有证据显示黑猩猩非常珍视肉类,将肉用作一种通货——一种从其他个体获取性、政治和物质好处的交易方式。[35]简而言之,食用更多的肉可能刺激智力新形式和社会复杂性的产生。
但是我们不能夸大肉在能人食谱中的重要性。我们不再相信,这些灵长目动物就像现代黑猩猩的某些群体一样是经常狩猎的。[36]对于能人牙齿的研究表明,即使肉类提供给他们特殊的补充食品,他们也主要靠水果和植物食品为生。另外,他们的石器与现代的食物采集民族相比,显然过于简单,这些石器在采集植物和食用腐肉时很管用,而在真正的狩猎中却几乎派不上什么用场。仔细检查能人遗址中发现的骨头上的砍斫痕迹,就可以发现他们切割动物的尸体,但是并不经常杀死动物,因为他们的切割痕迹通常是在别的动物的牙齿痕迹之上。他们可能杀死过小动物,但是或许他们以自然死亡或是被其他动物杀死的大型动物腐肉为食。
解剖学的研究同样揭示了能人并不完全是两足直立行走的,他们可能也有一些时间栖息在树上。所以我们可以想象由5—30个能人个体组成的群体在白天分散采集植物,就像现代的灵长目动物或是南方古猿,可能到了晚间他们就一起到树上藏身。尽管以腐肉为食对他们更重要,并且在陆地上生活的时间也更长,但是他们更加喜欢类似南方古猿那样的生态龛。
总而言之,并没有明显证据证明,路易斯·利基最早遭遇到的能人存在他所断言的智力和社会复杂性上的突飞猛进。
更大的脑量和活动范围:匠人和直立人
能人和其他几个种类,包括粗壮型的南方古猿(傍人属)同时居住在东非。事实上,人亚科原人以一种在早期新的适应——譬如直立行走——历史中相当普遍的范型,在其早期历史上显示出了极大的多样性。在能人的同一时期大概还生活着6种或者更多的人亚科原人。
大约180万年前,在地质学时间范围内由上新世到更新世的跃迁过程中,出现了一种新的人亚科原人,现代人类学家称之为直立人(Homo erectus)或是匠人(Homo ergaster)。[37]一个保存十分完好的匠人样本是1984年在非洲肯尼亚的尼奥科托姆(Nariokotome)发现的,其年代距今大约180万年。这个化石被称为“图尔卡纳男孩”(Turkana Boy),是所有古人类化石中最完整的。图尔卡纳男孩死时仍是个孩子,但是他已经有1.5米高和大约880毫升的头脑,几乎比大多数能人个体的脑量大了1/3。[38]
到了100万年前,在人类一次更壮观的辐射中,各种直立人/匠人取代了其他一切形式的人亚科原人。匠人个体比能人个体高大,并有着更大的脑容量,范围在850毫升到1000毫升之间。这使他们接近于现代人类的脑容量的标准。还有其他一些标志说明他们明显更接近于现代人类。从大概150万年前,他们开始制造一种新类型的石器,被称为阿舍利文化期(Acheulian)手斧,制造这种工具比奥杜韦文化期的工具需要更多成熟的智力因素。打造出来的形状比奥杜韦文化时期的砍砸器更精确、更优美。它们的每一个面都经过打造,形成一个梨形的“斧子”,通常至少有两个切割边缘。有时阿舍利文化的石器用骨锤打造出精美的边缘。一些匠人也学会了使用火。这为他们,特别是穴居生活提供了极好的保护,同时还可以烹饪肉类,使其更为柔软、清洁。不过,即使他们能够使用火,也不是系统性的。例如,没有证据证明他们使用炉膛。[39]
匠人的语言能力似乎要比能人强,但是又很难说到底强多少。较大的前脑表明理解和处理符号的能力有所增强,而喉处于喉咙较低位置,使得发音更加容易;因此,与手势交流相比,有声交流的重要性有所增强。但是,依然很少有证据能够直接证明他们拥有在现代人类的化石证据中非常明显的、丰富的符号活动能力,因此看来即使存在某些形式的符号交流,但是还并未在匠人的行为或意识中产生革命性的影响。[40]史蒂文·米森(Steven Mithen)有个有趣的观点,他说个别匠人可能主要在社交场合中使用他们的语言能力。[41]没有证据表明语言被用来解决技术问题,因为匠人的阿舍利文化期石斧自从出现后的100万年中几乎就没有什么改变。另外,尽管匠人通常所吃的食物中肉类含量要比他们的近亲能人所吃的更多,他们似乎不太可能像现代的食物采集族群那样从事系统的狩猎活动。
这些物种的行为流动性有所增长,其中一个最重要的标志就是,它们是最早迁移出东非,然后又走出了非洲,进入欧亚大陆的人亚科原人。大约在70万年前,直立人共同体居住在亚洲南部的部分地区,甚至进入到冰川时代的欧洲。直立人的化石遗存最早于1891年在印度尼西亚发现,而以20世纪20年代今北京郊外周口店山洞的发现最为著名。总而言之,直立人较之能人开发了更宽广的生态龛——既指生态范围上的“宽广”,也指地理范围上的“宽广”。特别是,他们显然已经掌握了在能人所不能适应的气候太冷或是季节性太强的地区生活。
一个物种所获得的生态龛的增长通常是以人口大发展为标志,我们可以合乎情理地假设,随着其所获得的生态龛数量的增长,人亚科原人的数量也随之增长。尽管我们并不知道任何一种早期人亚科原人的数量,但是他们的数量可能与20世纪前大猿的数量相似。可能在一段时间里有数万或是好几十万个人亚科原人,当他们迁移到非洲的西部、北部和欧亚大陆南部时,他们的数量就大为增长了。但是即便是直立人,我们也还没有发现其数量长期增长的证据。所以我们不应该夸大其走出非洲的重要性。在欧亚大陆南部,直立人进入的环境较之东非热带草原更具有季节性,但是其他方面十分相似。许多其他哺乳动物包括早期的人猿总科动物在内早就进行过类似的迁移。最后,令人吃惊的是,直立人并没有设法到欧亚大陆北部寒冷的中心地带居住。[42]也没有证据显示他们越过海洋到达澳大利亚和巴布亚新几内亚。
过去100万年中前人类的人亚科原人
在过去100万年中,非洲和欧亚大陆各地出现了若干类型的人亚科原人。无论在哪个地方,这些物种的大脑都迅速发展。最终,许多物种的脑容量都达到1300毫升,这已使他们归入现代人类脑容量范围之内。从大约20万年前开始,在经过很长一段几乎毫无技术变化的时期,一种新的石器制作技术出现了:被称为勒瓦娄哇文化(Levallois)或穆斯特(Mousterian)文化工具。在这些工具中,有一种形似龟壳的石核,它是经过精确计算,只一次就敲掉若干石片而成形的。我们可以假设,一套更丰富的工具总是同新的生态龛的开发联系在一起的。
为什么人亚科原人的大脑会如此迅速成长?要以充分的理由解释大脑成长比看上去要困难得多,因为脑容量大的动物非常稀有。可以论证,现代人类的大脑是我们所知的最复杂的物体。爱德华·威尔逊论证到,人类大脑的进化是地球生命历史的四大转折点之一。[43]每个人的大脑包含有大约1000亿个神经细胞,就和一个普通银河系里的恒星数量一样多。它们相互联结(平均每个神经细胞可能要与100个其他神经细胞相联),形成了惊人的复杂网络,包含长达9.6万千米的连锁。这样一个结构能够进行平行的运算。这就意味着尽管每一次运算可能比现代计算机速度慢一些,但是在一个特定时刻所能进行的运算数量的总和却要大很多。一台快速的现代计算机可在1秒钟内完成10亿次运算,而一只苍蝇即使在休息状态中,其大脑能够处理的次数也至少是计算机的100倍![44]当然,要进化成为如此强大的生物计算机必须具备一种很好的达尔文进化论的推动力。
但是尽管这个论证在直观上似乎合理,它仍有一个很严重的问题。如果大脑是如此明显“适应性的”,为什么只有少数物种真正进化出一个与其身体相比更大的大脑呢?麻烦在于维持大脑相当昂贵。人类大脑使用的能量是维持一个人全身所需能量的20%,但是其重量只占到身体重量的3%。生育一个大头的婴儿也是困难而危险的,对于直立行走的物种来说更是如此,因为直立行走需要一个窄臀而不是宽臀。换句话说,大脑增长是一个前途未卜的进化冒险。所以我们不能仅仅断言,因为具有显而易见的优势所以大脑就会进化,而是必须去寻找更加特殊的解释。
一个可能的解释是大脑为住在开阔空地的物种提供了一种很好的辐射。这个解释并不像它听上去那么轻率。但是还应该有更好、更妙的答案。可能有回馈循环,其中包括鲍德温的进化形式。某个领域(包括遗传上的或行为上的)变化可能会引起其他领域的变化,这就生成了新的选择压力来加强最初的变化。使用工具和大脑容量之间的关系可能就是我们所看到的这样的循环。
第二,社会性与大脑容量互为循环,紧密联系。即使在黑猩猩中,也可以看到准确思考社会关系的能力可以提高个体繁殖的机会。这样的过程可以建立起相对快速的回馈循环,因为社交能力较强的个体比较频繁地配对,生产较多的后代,而这些后代反过来又拥有了较强的社交和政治的能力。最后,这样的过程能够刺激最能进行复杂的社会性思考的那部分物种的大脑发展。[45]然而,大脑更大也使得生育成为一个更加疼痛和困难的过程。在某些时期,这个问题可以通过改变儿童的发育速度而得以解决。人亚科原人的孩子是在成熟阶段的早期出生的。但是这个解决方案意味着婴儿的自生能力变得越来越差,需要父母更多的抚养。这样就增加了母亲的重要性,母亲被一个男女组成的支持性的社会团体所包围。这种转变是和这样一个事实有关:与许多其他(除了猩猩以外)的大猿不同,人类丧失了发情期;因此,他们即使在不可能怀孕的时候也能够发生性行为。这种性和繁殖的部分分离加强了男女之间的配对关系,由此提高了男性在养育子女中的作用,这个变化也许和人类两性异形的衰退有联系。[46]无论这些复杂过程的细节到底如何(考古学的记录相当含糊,无法确定),人亚科原人随着他们脑容量增加而变得更加社会化。但是正如我们所知,在更大、更复杂的社会共同体中生活需要复杂的社会技能,社会技能最强的个体最容易找到配偶。这种类型的回馈循环——增长的脑容量刺激了社会复杂性的增长,而社会复杂性的增长又鼓励了脑容量的再次扩张——可以解释为什么在人亚科原人进化的某些阶段,人类的大脑(特别是额叶前部皮层)发展得特别迅速。[47]
另一种可能性是,大脑的成长是人亚科原人发展计划中一些微小变化的副产品。正如我们所见,幼态持续——或者物种的进化类似于它们所进化来的物种的幼年形式——的发生是由于主宰发展速度和时间的遗传密码的微小重组所致,因而一个物种除了性成熟之外的大多数特征都发展缓慢。于是,成年人类脸部平坦,相对而言毛发较少。黑猩猩也有这些特质,但只是在它们的青年期里。随着年龄的增长,它们口鼻向外突出,并且变得多毛。最重要的是,现代人类保持了典型的黑猩猩幼年时期大脑成长的速度,但是现代人类将这种成长的速度继续维持了更长的时间。这就意味着他们发育出更大的大脑,并且把这种幼年时期快速学习的脚步持续了更长一段时间。以此方式,控制发展过程的基因的小小改变可能对成年的幼态持续物种产生巨大的影响。
最后一种可能性是大脑的迅速成长与比较成熟的语言形式的进化有关。随着工具的使用,语言能力可能与大脑的能力发生了紧密的相互关系,赋予那些拥有较大脑容量的个体一个重要的达尔文进化论的优势。它将以进化回馈循环的方式促进更大脑容量的进化。我们将在下一章更仔细地探讨这一论证。
无论何种原因,我们知道人亚科原人的大脑从50万年前开始迅速成长。这些变化可以证明智力能力明显提高,可能也使语言能力明显提高。但是,令人沮丧的是,并没有证据表明,在人亚科原人生活方式中有革命性的变化。这些后期的人亚科原人中最著名的是尼安德特人(Neanderthal)。最早的尼安德特人化石是1856年在德国的尼安德河谷发现的。尽管长期以来尼安德特人(学名Homo sapiens neanderthalensis)被划分为与现代人类相同的种,但是最近用尼安德特人化石中残留的脱氧核糖核酸进行的基因测试表明,人类与尼安德特人的进化可能远在70万年到55万年前就分道扬镳了。[48]
最早出现于考古学记录中的尼安德特人生活在13万年前,他们在近至25000年的化石记录中消失。他们的脑容量和现代人类一样大,甚至可能比现代人类更大,但是他们的身体要比现代人类更结实、更矮壮。他们显然有狩猎的能力,这种能力使他们能够占据了此前从未有任何早期人亚科原人居住过的冰川时期的地区,例如,现在乌克兰和俄罗斯南部的部分地区。然而,他们的狩猎方法与现代的食物采集族群,甚至与旧石器时代晚期的人类相比,并不特别有效,也没有形成体系。他们的石器通常被称为穆斯特文化,比起直立人的石器要来得更为复杂,但是比起现代人类的石器则缺乏多样性和精确性。尼安德特人有艺术或是葬礼的迹象,二者都标志着符号交流日益增加(但是证据尚不明确)。同时也几乎没有迹象表明存在更大的社会复杂性。就像早期的人亚科原人,尼安德特人似乎仍主要以简单家庭为单位过着群体生活,这种方式限制了他们相互之间的交流。也没有证据证明尼安德特人对这个星球产生了像现代人类那样重要的影响。
本章小结
这是一个令人沮丧的结论。我们已经看到,在地球的历史中,现代人类的进化是一件革命性的事件。我们也看到,经过几百万年,所有的现代人类的基本要素都聚集了起来。人亚科原人进化出了较大的脑容量,这使得他们增强了行为的适应性,可能符号语言能力也开始加强。他们比其他灵长目动物学会以更复杂的方式使用工具,这使他们有机会获得更丰富的食物。这些变化综合在一起,显然使得直立人比其他相近的种拓展了更为广阔的居住范围。但是并没有清晰的化石记录能够证明,即使距今25万年前的晚期人亚科原人物种的行为方式存在任何革命性的变化。我们依然没有离开自然选择的领域,在这个领域,遗传变化超过了文化变化。很难想象人亚科原人早期的种如何能像我们这个种那样改变这个世界。这对于尼安德特人,这个在遗传上与我们惊人地相似的种、有着和我们一样甚至比我们更大的大脑的种而言,情况也是如此。那么,什么是现代人类和人类历史的革命呢?在这一章所描述到的变化以何种方式为他们革命性的生态影响做了铺垫?下一章中将提供一些尝试性的答案。
延伸阅读
关于人类进化有许多优秀的大众读物,但是这一领域的变化相当迅速,所以这些读物也更新得很快。其中,罗杰·卢因的《人类进化》(1999年,第4版)是最好的读物之一,而史蒂文·琼斯等人编著的《剑桥百科全书·人类进化》(1992年)则是一本极佳的参考书。这一领域的两位重要人物,理查德·利基和唐纳德·约翰逊都写过相关主题的著作[利基的《人类起源》(1994年);约翰森与艾迪合著的《露西》(1981年)]。贾雷德·戴蒙德的《第三种黑猩猩》(1991年)就该领域做了一次出色的概述,保罗·埃利希的《人类的本性》(2000年)是近期另一次全面的概览。其他全面的概览还包括戈兰·布伦哈特编的《图说人类历史》(5卷本,1993—1994年);布莱恩·费根的《地球上的人们》(2001年,第10版)是一本被广泛使用的教科书;罗伯特·福利的《人类以前的人类》(1995年);伊安·塔特萨尔(Ian Tattersall)的《成为人类:进化与人类的独特性》(1998年);罗伯特·温克(Robert Wenke)的《史前史的范型:人类的前3000年》(1990年,第3版);以及彼得·博古茨基的《人类社会的起源》(1999年)。克莱夫·甘布尔的《时钟》(1995年)是对旧石器时代的最好的全面纵览之一。在意识和思想的进化方面,史蒂文·米森的《头脑的史前史》(1996年)、泰伦斯·迪肯的《使用符号的种群》(1997年)、史蒂文·平克的《语言本能》(1994年)和《心灵如何工作》(1997年)、威廉·卡尔文的《思维的提升》(1991年)和《大脑如何思维》(1998年),以及尼古拉斯·汉弗莱的《心灵的历史》(1992年)都是很有价值的著作,尽管仍有许多领域需要继续思索。克雷格·斯坦福(Craig Stanford)的《猎猿》(1999年)和《重要的他者》(2001年)以敏锐的洞察力提出,现代灵长目动物学必须为人类进化的历史提供怎样的依据。在《非零》(2000年)中,罗伯特·赖特论述了非零和博弈在人类历史中所起到的重要作用。
[1] 例证参见约翰·梅纳德·史密斯和厄尔什·绍特马里:《生命的起源:从生命诞生到语言起源》(牛津:牛津大学出版社,1999年)。
[2] 最优秀的人类能量利用简史参见斯米尔:《世界历史中的能量》(布尔德:韦斯特韦尔出版社,1994年)。
[3] 保罗·埃利希:《自然界机械论》(纽约:西蒙和舒斯特出版社,1986年),第287页。陆地上人类适当的NPP数据来自于I. G.西蒙斯:《地球外貌的变化:文化、环境和历史》(牛津:布莱克韦尔出版社,1996年,第2版),第361页,他采用的是J. M.戴蒙德(J. M. Diamond):《人类对世界资源的利用》,载《自然》,1987年8月6日,第479—480页。
[4] 《世界资源报告2000—2001:人与生态系统:正在破碎的生命之网》(华盛顿特区:世界资源研究所,2000年),第246、248页。
[5] 参见理查德·利基和罗杰·卢因:《第六次生物灭绝:生命的模式和人类的未来》(纽约:达布迪出版社,1995年)。
[6] A. J.麦克迈克尔:《危险的地球:全球环境变化和人类的健康》(剑桥:剑桥大学出版社,1993年),第33页。
[7] 梅纳德·史密斯和厄尔什·绍特马里提供了如下的对生物领域的复杂体的新形式的描述:“那些在转变之前有能力独立复制的实体毕竟只能够作为一个较大的整体的一部分进行复制。”(《生命的起源》,第19页。)
[8] 克雷格·斯坦福:《猎猿:食肉与人类行为的起源》(普林斯顿:普林斯顿大学出版社,1999年),第28—29页。
[9] 约翰·密尔斯:《农业起源的全球透视》,参见迈克尔·阿达斯(Michael Adas):《古典史中的农业乡村社会》(费城:天普大学出版社,2001年),第65页。关于人类历史中非零和博弈(即在游戏中分享共同成功而不是竞争)的重要性的讨论参见罗伯特·赖特:《非零:人类命运的逻辑》(纽约:兰登书屋,2000年)。
[10] 麦克迈克尔:《危险的地球》,第34页。
[11] 德里克·比克顿:《语言和种族》(芝加哥:芝加哥大学出版社,1990年),第157页;引自威廉·卡尔文:《大脑如何思维——智力演化的今昔》(伦敦:菲尼克斯出版社,1998年),第82页。
[12] 泰伦斯·W.迪肯:《使用符号的种群:语言与脑的联合进化》(哈蒙斯沃思:企鹅出版社,1997年),第397页。
[13] 克里斯·斯特林格和罗宾·麦凯:《走出非洲》(伦敦:凯普出版社,1996年),第150页。
[14] 与其他许多伟大的科学文章相比,文森特·萨里奇和艾伦·威尔逊的这篇论文题目要缺乏诗意得多,《人类进化的免疫学时间比例》,刊登在1967年12月1日《科学》,第1200—1203页。
[15] “分子分类学”的局限性参见罗杰·卢因:《插图版人类进化》(牛津:布莱克韦尔出版社,1999年,第4版),第41—45页。
[16] 最近对早期人亚科原人遗存的发现意味着古生物学者可能在未来有能力用更多权威的证据来描述这一共同祖先。当然,还存在着诸多值得思索的问题,参见罗杰·卢因:《人类进化》,第84—85页。
[17] 关于环境和人类进化,参见卡尔文的《大脑如何思维》的简明摘要,第69—81页,以及威廉·卡尔文:《思维的上升:冰川时期的气候和智慧的进化》(纽约:矮脚鸡出版社,1991年)。
[18] 克雷格·斯坦福最近的两本书:《猎猿》和《重要的他者:猿——人连续统一体和寻求人的本性》(纽约:基本图书出版社,2001年)对这一领域的最近研究成果进行了概述。
[19] 我用了食物采集民族(foragers)这一术语,而不是通常的表达方式狩猎和食物采集民族,因为日益明显的是,在这样的社会里采集植物食物通常更为重要,至少从食物结构来看,要比肉类采集更为重要。这一术语是理查·李在其颇有影响的对桑人(!Kung San)的研究著作《桑人:一个食物采集社会的男人、妇女和工作》(剑桥:剑桥大学出版社,1979年);对采集技术的介绍参见艾伦·W.约翰逊(Allen W. Johnson)和蒂莫西·厄尔(Timothy Earle):《人类社会的进化:从食物采集团体到农耕国家》(斯坦福:斯坦福大学出版社,2000年),第3章。
[20] 参见罗伯特·弗里:《现代合成的阴影?近50年来古人类学研究的选择性观点》,载《进化人类学》第10卷,第1期(2001年),第5—15页,该文概述了关注于适应辐射的人亚科原人的历史。
[21] 参见罗杰·卢因:《人类进化》,第10章。
[22] 罗杰·卢因:《人类进化》,第55页。
[23] 安·吉本斯:《寻找最早的亚科原人》,载《科学》,2002年2月15日,第1214—1219页,这一组物种的学名是原始人图根种。
[24] 约翰尼斯·哈尔·塞拉西(Yohannes Halle Selassie):《中新世晚期埃塞俄比亚中阿瓦什的亚科原人》,载《自然》,2001年7月12日,第178—181页。
[25] 两足行走并不仅仅局限于人亚科原人,近来在地中海岛屿发现的一个900万年前的类似于猿的生物化石,它似乎就是两足行走的(斯坦福:《猎猿》,第220页)。
[26] 参见罗杰·卢因:《人类进化》第17章“两足动物的起源”。
[27] 里夫斯、罗斯奈、伊夫·科庞和多米尼克·西莫内:《起源:宇宙、地球和人类》(纽约:阿卡德出版社,1998年),第152—156页(科本斯的章节);罗杰·卢因:《人类进化》,第108—109页。
[28] 里夫斯、罗斯奈、伊夫斯·科庞和多米尼克·西莫内:《起源》,第156页(科本斯的章节)。
[29] 在这章及下两章里,年代将表示为如考古学家们常用的“距今”。严格地说,“今”在放射测年代技术中往往指的是20世纪50年代,但这其间的差距可以忽略不计。
[30] 露西的发现使得早期人亚科原人的进化令人着魔,参见唐·约翰逊和詹姆斯·施里夫:《露西的孩子:发现人类的祖先》(哈蒙斯沃思:企鹅出版社,1989年)。
[31] 参见斯坦福:《猎猿》。
[32] 关于人亚科原人两性异形的重要意义及其论据,参见沃尔特·洛特纳格:《两性异形:从比较与进化的视角》,载戈兰·布伦哈特编:《图说人类历史》第1卷:《最早的人类:人类起源及其至公元前1万年的历史》(旧金山:哈珀旧金山出版社,1993年),第41页。
[33] 伊安·塔特萨尔:《成为人类:进化与人类的独特性》(纽约:哈考特·布赖斯出版社,1998年),第133—134页。
[34] 近来,学界有人提出能人有两个种:脑量和体型都较大的卢多尔夫人和相对较小但颚和牙齿都更为现代的能人(参见罗杰·卢因:《人类进化》,第124页)。
[35] 在《猎猿》里,克雷格·斯坦福通过探索原始社会肉食在营养和社会方面的重要性,提出了一个关于“猎人”假设的修改后的观点。
[36] 有关黑猩猩的狩猎,参见斯坦福:《猎猿》。
[37] 通常称这一种的非洲样本为匠人,而非洲以外的样本为直立人。文中只有在特别讨论生活在非洲以外的样本时用“直立人”。
[38] 对匠人的简短精彩说明可参见保罗·埃利希:《人类的本性:基因、文化和人类期望》(华盛顿特区:亚伦出版社,2000年),第92—96页。
[39] 约翰·古德斯布洛姆曾提出使用火标志着人类历史中一个最基本的跃迁,参见约翰·古德斯布洛姆:《火与文明》(哈蒙斯沃思:亚伦·莱恩出版社,1992年)。
[40] 迪肯:《使用符号的种群》,第358页。
[41] 史蒂文·米森:《心灵的史前史:寻找艺术、宗教和科学的起源》(伦敦:泰晤士与哈得孙出版社,1996年),第179页及以下。
[42] 大卫·克里斯蒂安的《俄罗斯、中亚和蒙古史》第1卷:《从史前到蒙古帝国时期的欧亚内陆史》(牛津:布莱克韦尔出版社,1998年)第2章中探究了欧亚大陆北部没有直立人的重要意义。
[43] 爱德华·威尔逊:《论统合:知识的融通》(伦敦:艾伯克斯出版社,1998年),第107页;该书第6章对人类大脑的现代理解做了最好的简介。威尔逊指出的另一个转折点是生命的起源、真核细胞的出现和多细胞生物体的出现。
[44] 罗伯特·卢因:《混沌边缘的生命》(伦敦:菲尼克斯出版社,1993年),第163页。
[45] 如此的回馈循环在尼古拉斯·汉弗莱的著作中提出过,概述参见卡尔文:《大脑如何思维》,第66—68页。
[46] 参见贾雷德·戴蒙德:《性趣探秘:人类性的进化》(伦敦:威登费尔德和尼科尔森出版社,1997年);唐纳·J.哈拉维(Donna J. Haraway):《类人猿、半机械人和妇女:自然的再发明》(纽约:鲁特罗奇出版社,1991年),第107页,试图解释现代人类社会中女性缺乏明显的发情期的重要性。
[47] 罗伯特·福利:《人类以前的人类》(牛津:布莱克韦尔出版社,1995年),第165—171页。
[48] 埃利希:《人类的本性》,第96页。