如何用神经生物学的语言解释上一章最后提出的观点呢?来自生物调节的证据表明:简单有机体无法意识到在演化上较古老的脑区中所发生的反应,也无法深思熟虑。举例来说,爬行动物的大脑只包括那些古老的脑结构,缺乏演化上较现代的脑区,但它们可以毫无困难地作出反应选择。一些观点认为反应选择是决策的初级形式,因为诸多研究都清楚地表明,起决定作用的不是一个可以觉知的“自我”,而是一组与决策相关的神经回路。
然而,现在已经广为人知的是,当社会性有机体面对复杂情境、不得不进行不确定性选择时,必须使用演化上更新的皮层区域,即新皮层。研究结果表明,新皮层的扩展与分工细化有关,也与复杂多变且无法预期的环境有关,新皮层的扩展能帮助个体更好地应对这些环境。就该观点而言,约翰·奥尔曼(John Allman)得出一项有价值的研究结果:排除体形的影响,以水果为食的猴子的新皮层面积大于以树叶为食的猴子(1)。以水果为食的猴子需要更为丰富的记忆,以便记住在何时何地可以寻觅到可食用的水果,避开贫瘠的果树和腐烂的水果。它们较大的新皮层为上述活动所需的事实性记忆能力提供了支持。
“低级、古老”的大脑结构和“高级、较新”的大脑结构之间有明显差异,这种差异使人们自然而然地对两者各自的功能形成了一种看似合理的观点。简单来说,就像一栋建筑,古老的大脑核心区域在地下室处理基本的生物调节功能,在此之上,新皮层则处理更高水平的心理功能。换句话说,“楼上”的皮层处理的是推理和意志,“楼下”的皮层下区域处理情绪以及与有机体本身有关的事务。
然而,就我看来,这个观点在描述理性决策的神经机制方面有失偏颇。首先,这个说法和我们在第一部分观察到的结论不符;其次,人的寿命在某些方面反映了推理的质量。而现在有结果表明,人的寿命不仅如期望的那样跟新皮层的大小有关系,同样也跟下丘脑皮层大小的增加有关,而下丘脑是“下层建筑”的主要部分(2)。传统观点认为理性器官是新皮层,但若没有生物调节的配合似乎不能运行,而生物调节一般认为是皮层下组织的功能。看上去,自然界不仅将理性器官构建在生物调节器官的上方,而且也要借助生物调节器官。我相信,驱力和本能之外的行为机制既使用了“楼上”的脑区,也使用了“楼下”的脑区,新皮层和旧皮层核心共同成为理性的神经基础。
这就产生了一个问题,理性和非理性过程各自与大脑的皮层和皮层下结构的关联程度几何呢?为了解决这个问题,我们下面要开始讨论情绪和感受,两者是生物调节过程的核心,并在理性和非理性过程之间、皮层和皮层下组织之间搭建了桥梁。
基本情绪与次级情绪
威廉·詹姆斯对人类心智有独特洞见,只有莎士比亚和弗洛伊德可与之媲美。大概在一个世纪以前,他对人类情绪和感受提出了一个惊人的假设。现在请思考一下他的论断:
如果我们想象某些强烈的情绪,然后试图从意识中剥离这些感受伴随的躯体症状,会发现什么都没有剩下,没有参与构建情绪的“心理成分”,只有一个冰冷的、中性的理性知觉。
紧接着,詹姆斯用强有力的论点来说明他的观点:
如果我们不能感受到心跳加快、呼吸急促、嘴唇颤抖、四肢无力、浑身起鸡皮疙瘩或五脏六腑的翻腾,那恐惧这种情绪还能剩下什么?同理,如果没有怒火攻心、满脸通红、鼻孔扩大、咬牙切齿、攥紧双拳想要动手,反而只有放松的肌肉、平静的呼吸和温柔的面孔,这能叫作愤怒吗?
詹姆斯的上述论断既超越了他所属的时代,也超越了我们所处的时代,我相信他抓住了情绪和感受机制的本质。遗憾的是,他余下部分的论述缺乏对情绪现象多样性和复杂性的探索,因而成了之后无休止的争议的源头,这对他来说也是不寻常的(4)。这里我无法对关于这一主题的诸多学术研究作出评价,请参阅其他学者的评论,如乔治·曼德勒(George Mandler)、保罗·埃克曼(Paul Ekman)、理查德·拉扎勒斯(Richard Lazarus)和罗伯特·扎伊翁茨(Robert Zajonc)。
人们对詹姆斯的观点提出的主要异议不在于他将情绪拆解为一个涉及躯体的过程,虽然这个观点也让评论家震惊,但异议却在于在他的观点中没有提及个体对触发情绪的场景的心理评估过程。他的观点可以解释人们在人生早期感受到的情绪,但是无法解释奥赛罗产生妒忌和愤怒的心理过程,也无法说明哈姆雷特在准备做自己厌恶的事之前在想些什么,同样无法给出麦克白夫人在促使丈夫暴怒行凶后还会得意忘形的原因。
詹姆斯的观点还有一个严重的问题,即没有提出与被情绪激发的躯体对应的感受生成的替代或补偿机制。在詹姆斯的观点中,躯体总是介入情绪过程中的。此外,詹姆斯也丝毫未提情绪在认知和行为中可能起到的作用。然而,正如我在导言中说的,情绪并非可有可无。情绪在交流中起着传递意义的作用,而且在下一章我会提到,情绪也在认知指导过程中发挥作用。
简单来说,詹姆斯阐述了一个基本机制。在该机制中,环境中的特定刺激通过先天的、固定的机制引起了特定躯体反应模式。在该过程中,没必要在反应出现之前去评估刺激的重要性。正如詹姆斯的经典名言所述:“所有能激发本能的物体都能诱发情绪。”
然而我们知道,作为一个社会性个体,在许多情况下,我们只有经过一个评估性的、主动的、非自发的心理过程,才会产生情绪。出于人类经验的特点,大量刺激和场景都和天生可以产生情绪的刺激绑定在一起了。对这些刺激和场景的反应可以被作为中介的心理评估过程过滤。也正因为这种思维性的、评估性的过滤过程,预制的先天情绪模式的范围和强度才有可调整的余地。事实上,存在一种对詹姆斯所说的基本情绪机制的调节过程。此外,似乎还存在其他神经过程,也可以产生这种被詹姆斯视为情绪过程本质的躯体感受。
我将在下面的篇幅中阐述我关于情绪和感受的观点。我将首先从个体历史的角度切入,厘清人生早期体验到的情绪和成人情绪的区别。人生早期体验到的情绪,已经由詹姆斯学派的“预组织机制”进行了充分的解释。而成人的情绪,是在早期情绪的基础上逐渐建立的。我决定把“早期”情绪称为基本情绪,“成人”情绪称为次级情绪。
|基本情绪
情绪反应在多大程度上是天生的呢?我想说的是,动物和人必然都不是天生就对熊或者鹰有恐惧情绪的,尽管一些动物和人天生就恐惧蛇和蜘蛛。我认为存在这样一种可能性,即当我们感知到外界或躯体内部某个刺激的特定特征时,就会产生某些先天性预组织的情绪反应。这些特征可以包括:体形,如大型动物;翼展,如飞翔的老鹰;动作类型,如爬行动物的动作;特定声音,如咆哮;某些躯体状态,如心脏病发作时感受到的痛苦。这些特征会被单独或被联合加工,并被边缘系统的一个部分,如杏仁核探测到。这一部分中的神经核团拥有一种倾向性表征,可以引发恐惧情绪出现时的躯体状态特征,并改变认知加工过程以适应恐惧的状态。接下来我们会看到大脑如何凭空“模拟”躯体状态,我们还会讨论认知改变如何发生。
请注意,为了引发相应的躯体反应,有机体甚至不需要辨认出熊、蛇、鹰,也不必知道是什么带来了痛苦。只要早期感觉皮层探测、分类出特定实体,如动物或其他物体的一个或多个关键特征,杏仁核便可以接收到关于特定实体存在的关联性信号。一个小婴儿把头藏起来并不是因为知道鹰是危险的,只是对一个有巨大翅膀的东西以特定速度飞过的场景作出了快速反应(见图7-1)。
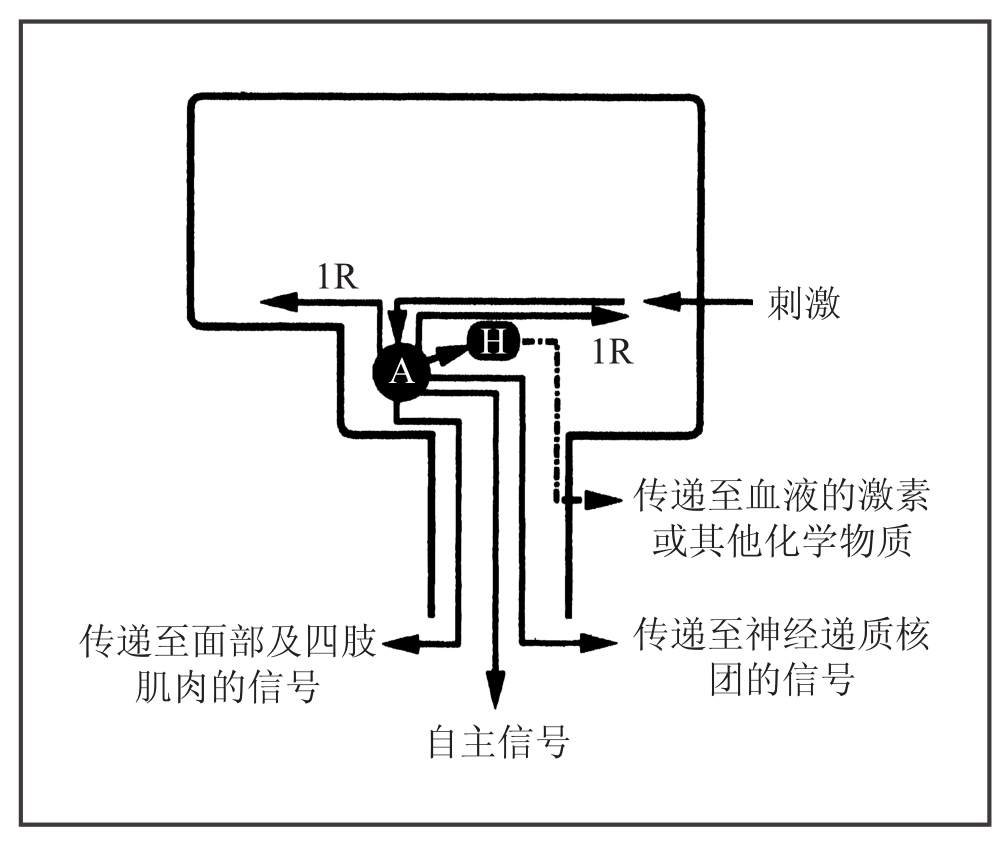
图7-1 基本情绪
黑色方框代表大脑和脑干。特定刺激激活杏仁核(A)后,产生了一系列反应:内部反应(IR),肌肉反应,内脏反应(自主信号),神经递质核团、下丘脑的反应(H)。下丘脑会释放激素和其他化学物质,并作用于血液循环。图中没有标注完成这些反应所需的其他一些脑结构。如表现情绪必须通过躯体姿势这类肌肉反应,这个过程需要基底神经节,即所谓的腹侧纹状体的参与。
情绪反应自身可以达成一些有利的目的,如快速躲避天敌,或向竞争对手展示愤怒。然而,这个过程不会因定义情绪的那些躯体变化的干预而停止。至少对于人类而言,这个循环过程会继续运作下去,下一步便是产生与激活这个过程的客体关联的情绪的感受,是意识到情绪化的有机体和该客体之间的关联。现在有人会问,为什么每个人都需要知道这其中的关联呢?如果已经存在某种自主的适应性反应方式,为什么还需要将问题复杂化并带入意识层面呢?答案是意识有利于扩大保护范围。考虑下面这个情况:如果你知道了某个可以导致恐惧的X,它可以是动物、物体或情境,对于X,你有两个应对策略。其一是天生的,你无法控制。此外,这个策略不仅限于应对X,其他大量的动物、物体和情境也都会激活该策略。其二是基于你自己的经验的,并且仅针对X。对于X的了解可以让你预先考虑并预测特定环境中X发生的概率,从而可以事先避开X,而不是只能在紧急情况下被动地对X的出现作出反应。
“感受”到自身情绪反应还有其他益处。如你可以一般化自身知识并决定对任何长得像X的物体警觉。当然了,如果你过度一般化,并且过度警觉,可能就会出现病理性恐惧,这当然不好。此外,在你第一次接触X期间,你可能已经发现了X行为的特别之处和潜在弱点。你可能会想在下次遭遇X时利用这个弱点,这也解释了你为什么需要“了解”这种关联。简而言之,感受你的情绪状态,就是说在意识层面上了解情绪,可以带来反应的灵活性,这种灵活性基于你同环境交互产生的特殊历史经验。尽管你需要一些先天机制来开启知识之旅,但情绪感受可以提供额外的东西。

基本情绪依赖于边缘系统回路,这个回路中的主要参与者是杏仁核和前扣带回。对动物和人的研究都发现杏仁核是预组织情绪的重要参与者。普里布拉姆(Pribram)、韦斯克兰茨、阿格里顿、帕辛厄姆所主持的多个动物实验都把杏仁核作为关注焦点,最近也可能是最广为人知的实验是由约瑟夫·勒杜主持进行的(5)。对该领域作出贡献的其他研究者包括E.T.罗尔斯(E.T. Rolls)、迈克尔·戴维斯(Michael Davis)和拉里·斯夸尔(Larry Squire)及其研究团队,尽管他们的研究目的主要是理解记忆,但同时也揭示了杏仁核与情绪的关系(6)。沃尔德·彭菲尔德、皮埃尔·格洛尔(Pierre Gloor),埃里克·哈尔格伦(Eric Halgren)在用电刺激技术研究癫痫患者的颞叶区域时也发现了杏仁核在情绪中的作用(7)。最近我的团队针对杏仁核的研究也支持了这一点。如果回顾相关研究历史,会发现杏仁核与情绪的关系的第一条证据来自海因里希·克吕维(Heinrich Kluver)和保罗·比西(Paul Bucy),他们发现,包含杏仁核的颞叶的部分区域被手术切除的患者,会有情感冷漠等一系列症状。关于前扣带回和情绪的关系,可参阅本书第4章以及相关研究文献(9)。
但是,这些基本情绪机制并不能解释所有种类的情绪行为。可以确定的是,它们是基本机制。然而,从个体发展的角度看,紧随其后的便是次级情绪。当个体开始进行感受,并在物体、场景的种类以及基本情绪之间建立系统性联系时,次级情绪就出现了。边缘系统中的结构不足以支持次级情绪的加工过程。上述过程涉及的网络必须扩展,并且还需要前额叶和躯体感觉皮层的参与。
|次级情绪
为了说明次级情绪的概念,我们用一个成人的经历作为例子。想象一下,你遇到了某个久未谋面的朋友,或被告知曾亲密共事的一个朋友突然去世了。无论在真实的情境中,还是在你此刻想象的情境中,你都能体验到情绪。在神经生物学层面,随着情绪产生,你的机体发生了什么?“体验到一个情绪”到底意味着什么?
如果你想象上述场景或类似场景时,恰巧我也在场,我也许可以进行一些观察。在上述场景中的关键方面,即遇到久未谋面的老友或同事去世的心理表象形成之后,躯体不同部分中的若干调整会带来躯体状态的改变。如果你遇到了一位老友,或在想象中遇到,你可能会心跳加速、皮肤发红,眼睛和嘴周围的肌肉会发生变化以展现出笑颜,其他部位的肌肉会放松下来。如果你听到了一位熟人去世的消息,你会胆战心惊、嘴唇发干、脸色苍白,部分内脏收缩,脖子和后背的肌肉变得紧张,面部肌肉呈现出悲伤的表情。次级情绪的发生过程可参见图7-2。
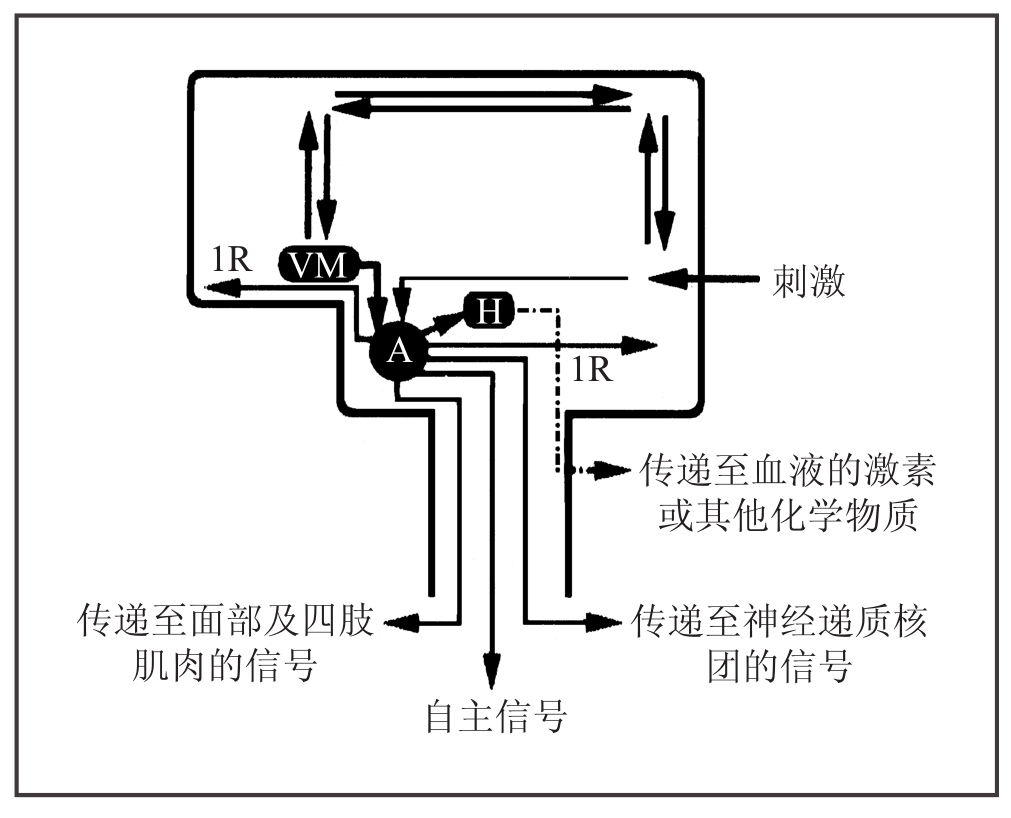
图7-2 次级情绪
刺激直接传递到杏仁核并且在思维过程中被分析,这进一步激活了前脑皮层(VM)。前脑皮层通过杏仁核(A)起作用。换句话说,次级情绪借助了基本情绪。此外,图中进行了简化,因为除了前脑皮层外的额叶皮层也被激活,但是我将主要部分勾勒在图中。请注意前脑皮层依赖杏仁核来表达其活动,也就是说,前者依附于后者。这种依赖关系说明了造物主工程学风格的思维方式。大自然充分利用旧结构、旧机制来创造新机制、获得新结果。
在上述任一情境中,内脏,如心脏、肺部、肝脏、皮肤;骨骼肌肉,如附着在骨骼上的肌肉;内分泌腺体,如垂体和肾上腺功能的相关参数都会发生变化。大量肽类调节剂从大脑释放到血液中,免疫系统也会快速调节。动脉壁平滑肌的基线活动水平提高,并导致血管收缩变细,造成面部苍白;或基线活动水平降低,使平滑肌放松,血管扩张,造成脸红。整体而言,这一系列改变勾勒出了对一系列平均状态背离的轮廓,这些平均状态与功能性平衡或内稳态机制有关。在这种状态下,躯体可以以最小的代价进行更快、更简单的调节。这一系列功能性平衡不是静态的,而是一个有着上限和下限的连续不断的动态过程。可以将此过程类比于河床。当你在河床上朝不同方向走时,你会看到有些地方凹进去,有些地方凸起来,此起彼伏。河床作为一个整体出现这些变化,但这些起伏变化都被河床的物理边界所限定,即容纳特定容量液体的边界。
在上段想象的情绪经历中,你躯体的许多部分都进入了新的状态,在其中发生了重要改变。那么,有机体是如何产生这些变化的呢?
1.这个过程始于意识,即对人或情境的深思熟虑。这种思考表现为由思维过程组织起来的心理表象,这些表象涉及了方方面面,包括你和特定人物的关系、你对当前情景的反思以及你和他人行为的结果。总的来说,这些表象是对你所参与事件内容的认知评估。其中一些表象是非语言性质的,如特定场景下特定人物的长相,而另一些表象是语言性质的,涉及特征、活动、名字等的词语或句子。这些表象的神经基础是一系列独立的拓扑性表征,这些表征位于不同的早期感觉皮层,如视觉、听觉或其他早期感觉皮层。这些表征是在分布于大量高级联合皮层上的倾向性表征的指导下构建起来的。
2.在非意识层面上,前额叶网络自发或本能地对来源于上述表象的信号作出反应。前额叶的反应来源于倾向性表征,这些倾向性表征丰富了个体知识,即在个体经验中,特定种类情景通常与何种情绪反应进行配对。换句话说,这些情绪反应是后天获得的而非先天的倾向性表征,尽管正如我们之前讨论的,获得性倾向也是在先天倾向的影响下获得的。后天获得的倾向性表征包含人生中类似关系的个体独特经验。你的经验跟他人的经验或多或少有所不同;这是你个人的经验。尽管在很大程度上,对每个人来说特定场景和情绪之间的匹配关系是相似的,但是个体经验赋予了每个个体独有的加工过程。总而言之,次级情绪依赖前额叶的获得性倾向性表征,这个过程独立于基本情绪所依赖的先天倾向性表征。接下来你也会发现,前者也需要通过后者来表达。
3.上一段提到的前额叶倾向性表征的反应可以非意识地、自发地、本能地把信号传递到杏仁核和前扣带回。前扣带回中的倾向性表征通过以下方式进行反应:(a)激发自主神经系统的核团以及经由末梢神经将信号传递给躯体,从而使内脏状态和触发该状态的情景种类保持一致;(b)传递信号给运动系统,从而使骨骼肌肉完成面部表情和躯体姿势这类情绪外貌的构建;(c)激发内分泌系统,其分泌的化学物质可以改变躯体和大脑状态;(d)以特定模式激发脑干和基底前脑的非特异性神经递质核团,使其释放神经递质到端脑的各个部位,即基底前脑和大脑皮层。这一系列纷繁复杂的动作是一个巨大的反应,它是变化的。这些多样化的反应针对整个有机体,对于健康的人而言,这真是个奇迹般的协调过程。
上述由(a)、(b)、(c)引发的变化作用于机体,并引起了“情绪性躯体状态”,这些变化的信号又被回传至边缘系统和躯体感觉系统。由(d)引发的变化则没有出现于躯体本身,而出现于一组负责躯体调节的脑干结构中。这些变化影响了认知过程的风格和效率,并构建了一个情绪反应的平行通路。我们将在下面对感受的讨论中,进一步说明(a)、(b)、(c)、(d)之间的差异。
现在很清楚的是,前额叶损伤的患者受损的情绪加工过程是第二种。这些患者无法生成与特定种类的情境和刺激所激发的表象相关的情绪,故而也无法产生感受。第9章中提到的临床观察和专门测试可以证实这一点。然而,这些前额叶损伤患者可以拥有基本情绪,这也是为什么乍一看这些患者的情绪反应是正常的,如有人在后面突然尖叫的话,他们也会被吓到,或遇到地震时他们也会惊慌。相反,边缘系统,如杏仁核、前扣带回损伤的患者通常在基本情绪和次级情绪两者上都有更广泛性的损伤,因而他们迟钝的情绪反应更容易被识别。
出于经济考虑,自然界没有为基本情绪和次级情绪的表达分配各自独立的机制。次级情绪的表达通道和基本情绪的表达通道是一样的。
我认为情绪的本质是一系列躯体状态的改变,这些躯体状态的改变通过神经细胞末梢在无数器官中被引发,并被特定脑系统所控制,这些大脑系统对应着特定实体或事件的思维内容。很多躯体状态的改变,如皮肤颜色、躯体姿势、面部表情等,是可以被外部观察者感知的。事实上,情绪(emotion)这个词的词源学解释就是从机体发出的向外的动作(movement out)。其他躯体状态的改变只能被情绪的主体本身感知到,但感知的更多是情绪本身而不是其本质。
总而言之,情绪是简单或复杂的心理评估过程与对这一过程的倾向性反应的组合,其中倾向性反应大部分指向躯体,结果是出现情绪性躯体状态。不过,其还可指向大脑本身,即脑干的神经递质核团,结果是引起额外的心理改变。请注意,这里我将构建情绪反应所需的,对所有改变的知觉过程排除在情绪之外。你将发现,我特意保留了“感受”一词用来描述对那些改变的感知。