
古代与现代的烤肉聚会:上图是画家笔下3.5万年前欧洲南部一个尼安德特人的营地,下图则为现代的类似场景(20世纪50年代的美国家庭)。我们是尼安德特人的后代吗?DNA的证据显示不是。
1856年8月,德国采石场工人在杜塞尔多夫(Du-sseldorf)外的尼安德山谷(Neander Valley)炸开一个石灰岩洞后,发现了一副不完整的骨骸。这些骨骸乍看之下像是属于一种已绝种的熊,这种熊的骨骸经常出现在洞穴里。但是当地一位教师发现,这副骨骸跟人类的亲缘关系近得多。这些骨头的主人身份成为争议的焦点,其中尤以头骨上厚厚的眉脊最令人费解。最奇特的说法是,这些骨骸属于一位受伤的哥萨克骑兵,他在拿破仑战争期间爬进这个洞穴后死亡。这个怪异的理论表明,这个可怜的家伙原先就有慢性病,老是皱着眉头,致使额部中间永久凹陷,且让眉脊特别突出。达尔文的《物种起源》出版后,引起各界对人类起源的热烈争论,4年后,也就是1863年时,这些在德国出土的骨骸主人被命名为尼安德特人(Homo neanderthalensis)。他们属于一种跟智人(Homo sapiens)不同但相似的物种。
虽然这副在德国出土的骨骸是最早被正式命名为尼安德特人的,但现在已经确认,更早在比利时和直布罗陀发现的骨骸也属于相同的物种。一个世纪后,更多的尼安德特人骨骸出土。现在的看法是,尼安德特人曾在欧洲、中东各处,以及北非部分地区定居,直到大约3万年前左右。尼安德特人之所以会有愚蠢笨拙的形象,大多是法国古生物学家布勒(Marcellin Boule)所造成的。他重建尼安德特人的材料取自法国的圣沙拜尔(La Chapelle-aux-Saints),而且是根据单一骨骸作出的,这副骨骸最后被发现是属于一位患有关节炎的老人的。事实上,尼安德特人的脑稍微比我们大一点(而且因为颅骨较平,所以形状跟我们不同)。埋葬地点的证据显示,尼安德特人的文化先进,已经有埋葬仪式,因此他们可能已经有来生的观念。
然而,尼安德特人引发的最大争议,主要在于他们跟我们的亲缘关系,而不在于他们有多聪明。我们是他们的后代吗?古生物学显示,现代人大约在尼安德特人消失时抵达欧洲。这两个群体之间是否曾经通婚,还是尼安德特人被现代人消灭了?由于这些事件发生在远古时代,现存证据又残缺不全(只有一些零星的骨骸),这类的争论还会延续许久,让学院派的古生物学家和人类学家乐此不疲。在尼安德特人典型的厚骨和现代人较轻的骨头之间,是否有另一种骨头存在?这类骨头有可能是这两个群体的混种,一个“失落的环节”。但它们也可能完全属于尼安德特人,只不过是某一位骨头特別轻的尼安德特人,或者它们有可能完全属于现代人,只不过是一位骨头特別厚的现代人。
令大家惊讶的是,这个争议已经由DNA解决:1997年从1856年出土的骨骸中取出的DNA,已有3万年之久。DNA的进化正是为了安全地保存信息,使其能代代相传,因此它的化学稳定性高,不会自动降解,也不会轻易和其他分子起作用。但它并非不会受到化学破坏。在死亡时,身体里的遗传数据会跟其他的成分一样,变得容易受许多降解者影响,这些降解者包括化学反应物质,以及能分解分子结构的酶。这些化学反应需要水,因此如果尸体脱水的速度够快,DNA就有可能保存下来。但是即使在理想的保存状况下,DNA分子可能顶多只能保存5万年。因此,要从已经有5万年之久、保存得并不好的尼安德特人骨骸里取出易读的DNA序列,实在是很不容易的事。
但是在德国慕尼黑大学,身材高大、说话简洁的瑞典人帕博(Svante Pǎǎbo)决定尝试解决这个问题。如果世上有人能做到,恐怕非他莫属。帕博是萃取所谓的“古代DNA”的先驱,先前他曾成功地取出埃及木乃伊和冰冻长毛象的DNA;1991年,有5000年历史的“冰人”因阿尔卑斯山冰河溶化而出土后,他也曾取过冰人的DNA。然而,尽管有这些了不起的成就,要钻到珍贵的尼安德特人遗骨里寻找完整的DNA,就算真的找得到,也是一项非常艰巨的任务。他的同事,考古学家史密兹(Ralf Schmitz)回忆道:“那就像是拿到割开《蒙娜丽莎的微笑》的许可证一样。”
帕博的研究生克林斯(Matthias Krings)负责执行这个计划。起初他不太乐观,但是在对遗骨的保存状态进行评估之后,初步的分析结果让他决定大胆做下去。克林斯的做法跟一般的预期不同,他不是在细胞核中搜寻DNA,而是把焦点放在微小的线粒体(mitochondria)上。线粒体位于细胞核外,遍布细胞各处,制造细胞所需要的能量。每个线粒体包含一小股DNA,长约1.66万个碱基对。由于每个细胞里有500个到1000个线粒体,而细胞核里只有两套基因组,所以克林斯知道就这些腐败的尼安德特人遗骨而言,在线粒体中找到完整序列的可能性,要比从细胞核中找到完整序列高得多。此外,由于线粒体DNA(简称mtDNA)长久以来是研究人类进化的主要对象之一,所以他有充分的现代人序列可供比较。
克林斯和帕博最担心的事情是污染。在定序古代DNA上有过许多例子,原先以为成功了,后来却发现这些古代样本曾遭到现代来源污染,得出的序列其实是错误的。我们每天都会有大量皮肤细胞脱落,许多DNA随之进入周遭环境,天晓得最后到了哪里。克林斯想用聚合酶连锁反应来扩增他想找的线粒体DNA片段,但聚合酶连锁反应非常敏感,即使只是一个分子都会引发作用,扩增它所遇到的任何DNA,无论这个DNA的来源是古代生物还是现代的活生物。万一尼安德特人的DNA降解得太厉害,以致聚合酶连锁反应无法对它发挥作用,却因为克林斯身上脱落的微小物质污染了古代样本,导致聚合酶连锁反应还是发生了,但扩增的却是污染源的DNA序列,那怎么办?如此一来,克林斯或许就得解释一下他和尼安德特人为什么刚好有相同的线粒体DNA序列。这对这位年轻人的顶头上司来说,肯定不是一个令人开心的结果,更不用说他的父母了。为了确保这种事不会发生,克林斯和帕博找了美国宾夕法尼亚州立大学的史东金(Mark Stoneking)实验室来依样重作他们的研究。那里也可能会有污染,但是至少不会是被克林斯的DNA污染,毕竟克林斯远在另一个大陆上。如果两个实验室从这个样本获得相同的结果,就可以合理假设他们找到了真正的尼安德特人序列。
克林斯在谈到他第一眼看到定序结果时说:“那是一种无法形容的兴奋感,好像有什么东西爬上我的背脊。”虽然,如同原先的忧虑,有些序列证明已被污染,但他也在一些其他的序列上看到很不可思议的结果:这些序列跟现代人的序列不同,但又具有令人好奇的相似处。把片段拼凑起来后,他重建出尼安德特人的线粒体DNA,总共有379个碱基对。但是当时宾州的结果还没出来。不过最后证明他们的序列是相同的,都有一模一样的379个碱基对。克林斯回忆说:“直到那时,我们才开香槟庆祝。”
这个尼安德特人序列跟现代人线粒体DNA序列之间的共同点,比它与黑猩猩之间多,这说明尼安德特人无疑是人类进化谱系中的一支。但在此同时,尼安德特人的序列,和克林斯拿来作比较的全部986个现代人线粒体DNA序列之间,也有惊人的差异。在这当中,即使是跟尼安德特人的序列最接近的现代人线粒体DNA序列,至少也有20个碱基对(占5%)是不同的。后来,另外两个尼安德特人(分别在俄罗斯西南部和克罗地亚出土)的线粒体DNA也被定序,而且如同预期,这两个序列跟首先被定序的那一个并不完全一样(尼安德特人之间自然也会有变异,如同现代人之间),但它们很类似。
基于这些遗传证据,我们可以下结论说:尼安德特人在人类及其亲戚的进化树上,的确占有一席之地,但他们那条分支跟现代人这条分支相距很远。如果3万年前他们在欧洲相遇时,尼安德特人和现代人真的曾经混种,尼安德特人的线粒体DNA应该会进入现代人的基因库。但是我们并没有看到这类输入的证据,这显示现代人消灭了尼安德特人,而不是跟他们混种。至于他们是在直接的冲突中消灭了尼安德特人,还是经由比较细腻的方式,就不是DNA可以告诉我们的了。
尼安德特人DNA的研究已经证明,我们和他们在遗传上明显不同。但就整体而言,有关人类进化的分子学研究似乎朝相反的方向发展:它们显示我们和自然界其余生物的遗传关系近得惊人。事实上,分子资料经常对以往有关人类起源的假设提出质疑,甚至推翻它们。
伟大的化学家泡令是从分子层面来研究进化的先驱。在20世纪60年代早期,他和分子进化学家祖克坎德尔(Emile Zuckerkandl)比较了数个物种之间相应蛋白质的氨基酸序列。当时正值蛋白质定序的初期阶段,数据自然相当有限。但是他们注意到一个惊人的模式:两个物种在进化上的亲缘关系愈近,他们相应的蛋白质序列愈相似。以血红素分子的一个蛋白质链为例,他们发现,在它全部141个氨基酸中,人类和黑猩猩之间只有一个氨基酸不同,但人类和马却有18个氨基酸不同。分子序列的数据反映出马和人类在进化上的分歧出现得比人和黑猩猩要早。找出隐藏在生物分子里的进化历史,现在是常见的做法,但在当时这却是具有争议的新概念。
研究进化的分子学方法取决于两个变量的相互关系:两个物种(或族群)在进化上开始分歧后的时间长短,以及他们之间的分子差异程度。这个“分子钟”(molecular clock)的逻辑很简单。为了方便说明,让我们假设有两对同卵双胞胎,一对男的,一对女的,每对双胞胎的基因完全相同。我们让两对双胞胎彼此婚配,然后让这两对夫妇各在一个荒岛上独居。从遗传学观点来看,这两个岛上的族群起初无法区分。现在让这两对夫妇及其后代在各自的荒岛上繁衍数百万年,等这段期间过去后,其中一个族群应该已经有了另一个族群没有的突变,反之亦然。由于突变的发生速率慢,再加上个体的基因组大,有许多位置都可能发生突变,所以这两个族群发生完全相同的突变几乎是不可能的事。因此,当我们为每对夫妇的后代定序DNA时,会发现这两对夫妇原本一模一样的基因组已经累积了许多差异,也就是说这两个族群已经有了遗传变异,而且互相分离的时间愈长,差异愈大。
但是我们要怎么从这个“分子钟”看出“时间”呢?换句话说,我们要怎么测量我们和自然界其余生物的遗传差异?在60年代晚期,早在定序DNA的时代来临前,加州大学伯克莱分校古怪的新西兰人威尔逊(Allan Wilson)和他的同事萨瑞奇(Vince Sarich)就已开始用泡令和祖克坎德尔的逻辑来研究人类和人类的近亲。当时,定序蛋白质仍然相当棘手费力,但是威尔逊和萨瑞奇却找到一个巧妙的快捷方式。
从免疫系统对外来蛋白质的反应强度,可以看出那个外来蛋白质跟被入侵的生物体之间有多少差异:如果它和这个生物体本身的蛋白质相当类似,则免疫反应也会相对较弱;但是,如果这两个蛋白质差异很大,免疫反应也会按比例变得较强。威尔逊和萨瑞奇从一个物种取出一种蛋白质,再测量它在其他物种体内引起的免疫反应,以此来比较反应强度。他们借此建立两个物种之间分子差异的指标,但是要让这个“分子钟”具有时间意义,他们必须实际测量。
化石证据显示,新旧世界的猴子(地球上两种最主要的猴群),大约在3000万年前从共同祖先开始产生分歧,因此威尔逊和萨瑞奇把新旧世界的猴子之间的免疫“距离”设定为3000万年。如此一来,人类和进化上关系最近的黑猩猩及大猩猩等大猿(great apes)之间的距离又是多少?1967年,两人发表了他们的估计,认为人类和大猿大约是在500万年前开始分道扬镳。他们的说法引起一阵哗然,因为古人类学家向来认为,人类和大猿早在2500万年前左右就已分离。传统理论坚持,人类和大猿间的差异,显然是在远远超过500万年以上的时间中进化而成的。许多人认为光凭这一点,就足以证明这个伯克莱团队新式的遗传学方法没有价值;他们宣称,遗传学家还是专心研究果蝇吧,把有关人类的研究留给人类学家!但是威尔逊和萨瑞奇还是撑了下去,而且后续的研究也证明,他们所定出的人类与猩猩在进化上的分歧时间相当精确。
威尔逊先前是用蛋白质来分析人类与猿猴在进化上的分歧时间,等到DNA技术成熟时,他将进一步的分析工作交给他指导的研究生玛丽—克莱尔·金(Mary-Claire King)。她在1975年写出的报告,是20世纪最杰出的科学论文之一。其实有很长一段时间,这个研究似乎不太可能有很好的结果,特别是从金的观点而言。她的研究进行得并不顺利,部分原因在于70年代初期伯克莱的反越战运动让人很难专心。金一度考虑到华府去为消费者保护健将奈德(Ralph Nader)工作,但幸好她听了威尔逊明智的建议:“如果每个做实验失败的人都不再搞科学,世上就不会有科学存在。”金决定继续她的研究。
金和威尔逊在比较黑猩猩和人类的基因组时,混合采取了数种方法,包括一种称为DNA杂合(DNA hybridization,亦称DNA配对法)的聪明方法。当DNA互补的两股合并形成双螺旋时,可以借由将它们加热至摄氏95度,把它们分离(此现象在分子遗传学上称为“融解”[melting])。如果这两股并非完全互补,也就是当其中一股有突变时,会是什么情形?结果发现这两股会在摄氏95度以下的温度就融解。至于这个温度有多低,则取决干这两股之间的差异程度:差异愈大,要使它们分离所需的热能就愈少。金和威尔逊用这个原则来比较人类和黑猩猩的DNA。两者的序列愈相似,其双螺旋的融点就会愈接近完全互补时的标准融点(摄氏95度)。相似程度的实验结果令人相当惊讶:金推论出人类和黑猩猩的DNA序列的差异程度仅在1%而已。事实上,人类和黑猩猩的相同程度,比黑猩猩和大猩猩之间还高。黑猩猩和大惺猩的差异程度大约是3%。
由于这个结果太过惊人,金和威尔逊觉得自己有必要为遗传进化速率(慢),和身体构造及行为进化速率(快)之间的明显差异提出解释。既然遗传变化这么少,为什么动物园里的黑猩猩和在玻璃窗外观看它们的人类之间,还会有这么大的差异?他们的看法是:大多数重要的进化改变,都是发生在控制基因启闭的DNA片段。如此一来,一个微小的遗传变化有可能造成重大的影响,例如这个小变化可能改变了基因表现的时机。换句话说,大自然可以借由调控相同的基因,让它们以不同的方式运作,从而创造出两种长相非常不同的生物。
接下来,威尔逊的伯克莱实验室又在1987年抛出一个最具震撼力的炸弹。他和同事肯恩(Rebecca Cann)利用DNA序列变异的模式,建立了整个人类的族谱。这是极少数能登上美国《新闻周刊》封面的科学成果之一。
如同克林斯在10年后对尼安德特人所作的分析,肯恩和威尔逊也是利用线粒体DNA。这么做的原因有好几个,但是,就像一般的情况,实际执行上的问题是最重要的考虑。在聚合酶连锁反应技术成为研究主流之前,要取得足够的DNA以研究某个基因或区段,是相当让人头疼的事。肯恩和威尔逊的研究必须分析147个样本,而不只是一个而已,因此他们需要用上所有找得到的DNA。人类组织样本的线粒体DNA比细胞核里的染色体DNA多很多,但是他们仍需要许多组织,才能取出足够的线粒体DNA。他们的解决方法是使用胎盘。医院在接生婴儿后通常会把胎盘丢掉,而它们正是线粒体DNA的丰富来源。肯恩和威尔逊需要做的是说服147位孕妇,把宝宝的胎盘捐出来供科学研究之用。事实上是146位,因为金很乐意捐出她女儿的胎盘。他们知道要使重建的人类族谱尽可能完整,他们所使用的组织就必须取自遗传多样性最大的捐赠者群中。在这方面,像大熔炉般的美国族群明显具有优点:他们不必远到非洲去找非洲人的DNA——非洲人的基因早已进入美国。但是他们得靠新几内亚及澳洲的合作伙伴去寻找愿意提供组织的原住民妇女(她们在美国的基因库中所占的比例并不高)。
我们的线粒体DNA都是从母亲继承而来。父亲的遗传物质包在精子的头部,其中并不包括线粒体的物质。精子的DNA被注入已含有母方线粒体的卵细胞内。因此,肯恩和威尔逊追踪的是人类母系的历史。由于线粒体仅在母系中传承,所以没有机会重组。所谓重组也就是染色体的臂部片段互相进行交换,所以突变可以从一个染色体移动到另一个染色体上去。在根据DNA序列的相似性重建族谱时,线粒体DNA没有重组的情形成为一大优点。如果两个序列有相同的突变,我们就知道它们一定有共同的祖先(也就是最早产生这个突变的生物)。如果有重组发生,这些谱系的突变就有可能是从刚刚发生的重组活动中获得的,所以拥有相同的突变并不一定代表有共同的祖先。因此,利用线粒体DNA来建立族谱的逻辑很简单:类似的序列(有许多相同突变的序列)表示亲缘关系近;有许多差异的序列表示亲缘关系比较远。从族谱的树状示意图(P185)来看,亲缘关系近的物种(由年代较近的共同祖先传承下来的物种)在“族谱树”上的距离较近;亲缘关系较远的物种分散得比较远,因为它们的共同祖先存在的年代比较久远。
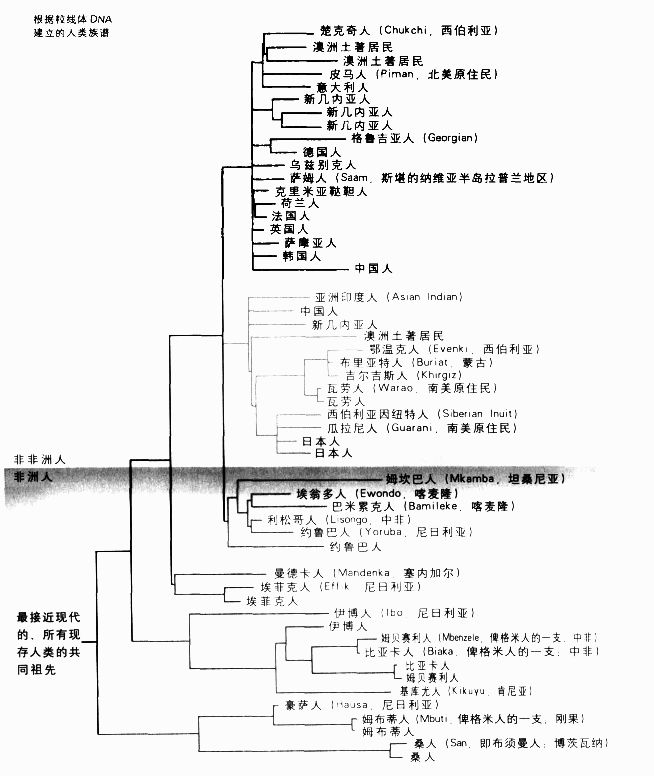
根据粒线体DNA建立的人类族谱
肯恩和威尔逊发现人类族谱有两个主要的分支,一支仅包含非洲的一些不同族群,但另一支则包含一些非洲族群和所有的其他族群。这表示现代人起源于非洲,也就是说我们所有人的共同祖先住在非洲。但这几乎称不上是新观念。达尔文在注意到亲缘关系跟我们最近的黑猩猩和大猩猩都是非洲原产的物种后,就曾推论人类也是在非洲进化的。在肯恩和威尔逊所建立的人类族谱上,最惊人和争议最大的一点,在于人类族谱最早可以追溯至多久远的年代。在对突变于进化中的累积速率作出一些简单的假设后,就可以计算人类族谱的年代,也就是一直往回追溯至我们的曾曾曾曾曾曾……祖母的年代。肯恩和威尔逊获得的估计值是15万年,换句话说,即使现存亲缘关系最远的人,也在15万年前有共同的祖先。

“线粒体夏娃”成了封面女郎:1988年1月11日出刊的美国《新闻周刊》以“寻找亚当和夏娃”的标题,报道威尔逊等科学家对人类起源的争议性理论。配合“非洲起源说”,亚当和夏娃以非洲人的面貌现身说法。
如同萨瑞奇和威尔逊20年前所获得的结果,许多人类学家这次也是群情哗然,不肯相信肯恩和威尔逊的研究结果。当时对人类的进化有个普遍的看法:我们的祖先大约在200万年前离开非洲,后来才在旧世界定居。根据这样的理论模型,人类族谱的长度应该是他们所推算的13倍。肯恩和威尔逊的看法被媒体称为“夏娃假说”(The Eve Hypothesis),或没那么夸张的“非洲起源说”(Out of Africa)。他们并没有否定在更古老的时代所发生的迁移活动,而是暗示现代人抵达欧洲后,取代了早期的“类人”(hominid)族群,这些类人的祖先在将近200万年前离开非洲来到了欧洲。200万年前从非洲向外扩散的直立人(Homo erectus)在旧世界四处迁徙,并在70万年前左右促成尼安德特人的兴起,因此尼安德特人其实是这些直立人在欧洲的后代。后来,顶多在大约15万年前,智人,或说现代人(他们也是直立人的后代,但是一直在非洲进化,没有离开过)和久远年代以前他们的直立人祖先一样,选择踏上了离开非洲的浩大旅程。我们在本章开头已经看到,尼安德特人没有和刚抵达欧洲的族群混种,这和现代人遇到直立人时的情形似乎相同。无论他们在什么地方相遇,现代人都会取代直立人。最后一批尼安德特人在2.9万年前左右消失,代表了最后一批非现代人的直立人后裔步入灭绝。
肯恩、威尔逊以及他们的研究伙伴,从此彻底改变了我们对人类历史的看法。
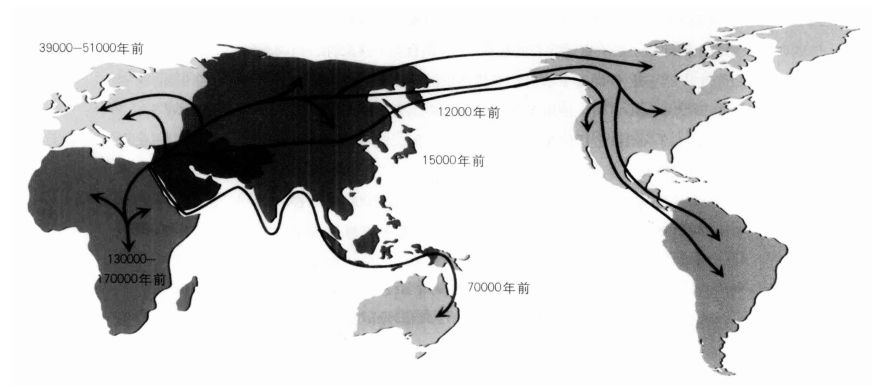
远离非洲:人类起源于非洲,并从非洲散播至世界各地。图中的移民年代是根据线粒体DNA的资料推估出来的结果。
后续的研究证实了肯恩和威尔逊的结论。新的研究结果大多来自卡瓦利-斯弗扎(Luigi Luca Cavalli-Sforza)在斯坦福大学的实验室,他是用遗传学方法来研究人类学问题的先驱。卡瓦利-斯弗扎出身意大利米兰的名门望族,对显微镜很着迷。他相当早熟,1938年16岁时就进入帕维亚大学(University of Pavia)医学院就读。他曾特别提及,“那是一个相当幸运的选择”。否则他的另一个选择是加入墨索里尼的军队。1951年我第一次遇到他时,卡瓦利-斯弗扎还是积极有为的细菌遗传学家。但是一位研究生闲聊间无心的一句话,让他从细菌遗传学转而研究人类遗传学。这位研究生曾接受成为神职人员的训练,偶然提及天主教会保存了超过三个世纪的详细婚姻记录。卡瓦利-斯弗扎发现这些记录可以提供丰富的研究资源,因而逐渐转向研究人类遗传学,他可能是极少数通过天主教教会找到职业专长的人类遗传学家之一。
卡瓦利-斯弗扎知道,只有由父传子的基因,亦即人类基因组中由父系传承下来的一些成分,才是肯恩和威尔逊对人类进化看法的最有力证明。如果能借由追踪父系,亦即不是走肯恩和威尔逊分析线粒体DNA的母系路径,而是走父系路径来获得相同的结论,就可以作为真正独立的确证。在基因组中,男性特有的成分自然就是Y染色体。根据定义,拥有Y染色体的人即是男性(要记得,Y染色体是男性自父方继承而来的,男性的精细胞不是含有X染色体,就是含有Y染色体;精子会跟永远只含有X染色体的卵细胞结合,因此精子便决定了我们的性别,XX组合会产生女性,而XY的组合是男性)。Y染色体握有了解男性遗传历史的关键。此外,由于重组只发生在成对的染色体之间,使用Y染色体不会碰上重组的问题,可以避免进化分析上的可怕陷阱。任何Y染色体都是独一无二的,绝不会有另一个Y染色体可以跟它交换遗传物质。
卡瓦利-斯弗扎的同事安德席尔(Peter Underhill)在2000年发表了一篇论文,造成轰动。安德席尔研究的是Y染色体,就像肯恩和威尔逊研究线粒体DNA—样,他的研究发现跟他们的极其类似。他再度发现人类族谱的根源在非洲,而且也再度证明了人类族谱很短:它不像人类学家原本所设想的,是一棵远古的高大树木,而是跟肯恩和威尔逊的分析结果一样,是一棵灌木,树龄只有15万年。
两批独立的资料,却对人类的历史有类似的发现,这是极为惊人的成果。在只研究一个区域,例如线粒体DNA时,所获得的结果即使很有意义,仍不能当做定论;这个模式所反映的,可能只是此特定区域的DNA历史所具有的特色,而不是某个重大历史事件对人类这个物种整体的影响。最重要的是:当“族谱树”的分支汇聚到一点——所谓的“趋同点”(convergence point),也就是我们研究的所有序列最近代的共同祖先、我们所有人的曾曾曾曾……祖母——这并不必然与人类历史上任何特定事件有关。虽然它有可能暗示现代人类的起源,或其他一些具有历史意义的人口学上的事件,但是,以人类历史的宏观角度而言,它也有可能只是显示出一些更为琐碎的事,例如过去自然选择对线粒体DNA的影响。然而,如果相同的改变模式可见于基因组中一个以上的区域,就很有可能代表真的找到了过去某个重要事件的遗传痕迹。
为了更加了解自然选择对遗传变异模式(以及人类族谱的整体年龄)的影响,让我们先假设下列的情况:在15万年前,有一个早期原始人类族群拥有过多的线粒体DNA序列,如同今日的人类,结果在这些序列中,有一个序列发生了有利的突变——自然选择偏好的突变。接着这个突变发生的频率会增加,直到许多世代后,这个种群的所有成员都有了这个突变为止。由于线粒体内不会发生DNA重组,线粒体DNA之间没有交换,自然选择过程会对有利突变首度出现的那个序列造成整体影响,这个种群的每一个成员最后都会拥有相同的线粒体DNA序列。因此等到自然选择的工作完成后,每一个个体都拥有了这个有利突变,而这个种群的线粒体DNA也不再有遗传变异。然而,随着时间逐渐推移,突变会再度发生,变异也再度增加,但是这些新的线粒体DNA序列终究都是源自于最早发生突变的那个序列:这就是族谱的趋同点,亦即所有序列最近代的共同祖先。这也正是肯恩和威尔逊发现的模式,但是在这个例子中,趋同点顶多只代表进化对线粒体DNA的微调(fine-tuning)。
这个模糊不清的问题一直纠缠着肯恩和威尔逊的研究结果:这究竟是进化的修补过程所造成的,还是在人类史前时代整体发展中的某一个重大事件所造成的?但是当安德席尔也在Y染色体上观察到类似的模式时,这个模糊性便消失了。两者的一致,强烈显示在15万年前,人类族群的确经历了一场剧烈的遗传变化,而且其剧烈程度已经足以同时对线粒体DNA和Y染色体造成影响。这当中涉及的现象即称为遗传瓶颈(genetic bottleneck),我们稍后将进一步讨论这点。
人口统计学上的因素对族谱有什么影响?任何系谱都是不同的血统相互消长的结果:随着时间流逝,有些会繁衍,有些会灭绝。以姓氏为例,假设在1000年前,某个偏远小岛上原本只有三种姓氏:Smith, Brown和Watson。再假设,当要把新生儿的名字抄到出生记录簿上时,偶尔会犯一些小错误(即突变)。这些错误很罕见,也很轻微,所以我们还能够分辨这个被写错的姓氏原本是哪一个:Browne显然是Brown的突变。现在假设过了1000年,到了现代,我们发现岛上的姓不是Brown,就是Browne, Bowne, Frown或Broun. Smith和Watson这两支已经灭绝,而Brown这一支成功繁衍下来(而且通过突变而变得多样化)。这代表发生了什么事?完全是运气,导致Smith和Watson这两支灭绝。例如,可能有一代的好几对Smith夫妇生的大多是女儿。假设岛上的姓氏是沿父系继承的(这是根据传统所作的假设,尽管现代已有其他的常规),生许多女儿有可能造成下一代中姓Smith的人减少。现在假设下一代的Smith也生育了太多女儿,人口效应就会再度增强,最后会完全没有了姓Smith的人。Watson这一支的情形也一样。
事实上,这种随机灭绝的情况在统计学上是不可避免的。然而,这种情况的发生速率通常很慢,以至于惟有在很长的时间后才会变得明显。不过有时瓶颈(人口规模锐减的时期)会大幅加速这个过程。在岛上的族群历史中,刚开始只有三对夫妇(六人),所以很可能在单单一代中便失去Smith和Watson这两支,因为他们只生女儿或无法生育的几率蛮高的。在大族群里,这种家系突然消失的情形不会发生;这在统计上是难以想像的事,如果一个族群里有许多Smith夫妇,很难想像他们全都只生育女儿或完全无法生育。惟有在经过许多世代后,家系逐渐缩小的效应才会慢慢浮现。事实上,我们假设的姓氏消失过程真的在南太平洋发生过:1789年著名的“叛舰喋血记”事件中,英国船舰“丰饶号”(Bounty)的6名叛变船员带着他们的13位塔希提岛新娘占领了皮特肯岛(Pitcairn Island)。在7个世代内,岛上的姓氏就减少到了3个。
现在回头来看我们虚拟族群中Brown, Browne, Bowne, Frown或Broun这几个姓氏。我们可以推论,他们全都源自起始三个谱系中的同一个谱系:Brown。那么人类线粒体DNA和Y染色体的数据所具有的含义,应该也不会让我们惊讶:15万年前,有许多不同的线粒体DNA序列和许多不同的Y染色体序列,但是今天的序列全都是其中一个线粒体DNA序列和其中一个Y染色体序列的后代。所有其他的序列都已经灭绝,大多数可能是在某个古代的“瓶颈事件”中消失的,这事件有可能是族群人口因瘟疫而锐减、气候改变或任何其他原因。但是无论我们早期历史中的这个灾难事件是什么,有一件事很清楚:在一段时间后,我们的祖先族群开始离开非洲,展开人类殖民全球的漫长旅程。
线粒体DNA和Y染色体的资料也证实了另一项有趣的发现,那就是南非的桑人 (San)在人类族谱中的位置。他们在人类族谱中的分支长度是最长的,也就是最古老的。这绝非显示他们比我们其余的人“原始”,每一个人类跟与我们亲缘关系最近的大猿之间,进化差距和分子差距都一样。如果我们沿着谱系追溯到黑猩猩和人类最后一个共同祖先,我的谱系会有大约500万年之久,而桑人的也一样。事实上,在这段漫长岁月的大多数时间里,我们这两个谱系都是相同的;一直到15万年前,桑人的谱系才跟其他人类的谱系分开。
(San)在人类族谱中的位置。他们在人类族谱中的分支长度是最长的,也就是最古老的。这绝非显示他们比我们其余的人“原始”,每一个人类跟与我们亲缘关系最近的大猿之间,进化差距和分子差距都一样。如果我们沿着谱系追溯到黑猩猩和人类最后一个共同祖先,我的谱系会有大约500万年之久,而桑人的也一样。事实上,在这段漫长岁月的大多数时间里,我们这两个谱系都是相同的;一直到15万年前,桑人的谱系才跟其他人类的谱系分开。
从遗传证据来看,桑人似乎在最初迁移至非洲南部和东部以后,在整个历史中都处于相对孤立的状态。从桑人罕有的(至少在我听来很稀罕)“喀嗒”(click)语言来看,社会语言学上的证据也足以支持这种推论。由于说班图语(Bantu)的族群在1500年前开始从中非西部向外扩张,所以桑人目前的分布范围极为有限。班图人的扩张迫使桑人迁至贫瘠的边陲地区,例如卡拉哈里沙漠(Kalahari Desert)。
从桑人相对比较稳定的历史来看,他们是否可以让我们窥知所有现代人的祖先是什么模样?有可能,但不必然。在过去的15万年间,桑人的谱系也可能发生过重大改变,与祖先不同了。即使想从桑人来推测我们早期祖先的生活方式,也是有疑问的:自从说班图语的族群在较晚的年代迁移到这里后,桑人退居沙漠一隅,而他们现在的生活方式是适应这种严酷环境的结果。2000年,我拜访卡拉哈里沙漠的一个桑人小区,体验了几天他们独特刺激的生活。我对他们惊人的实用主义生活方式感到相当震撼,他们以有效率又实际的方式执行眼前的所有工作,即使那些不是他们平常会做的事也一样,例如修理漏气的轮胎。那时我真希望我有更多适应能力可以跟他们一样好的同事!从遗传观点来看,纵使他们跟我的遗传相差十万八千里,但我们竟然在心性上这么相似,实在令我震惊。
桑人独特的遗传与文化特色不久就会消失。卡拉哈里沙漠的年轻人不怎么想跟他们游牧的父母一样,过着简单的采集/狩猎生活。例如我去拜访桑人族群时,他们表演了一场传统的“神灵附体之舞”(trance dance),但是年轻的族人显然对长老们古怪的舞姿感到困窘。他们迟早会搬离自己的小区,跟其他族群通婚。
事实上,桑人和其他族群逐渐混合的趋势已见诸历史。例如南非共和国黑人领袖曼德拉所属的科萨部落(Xhosa)就混合了班图人与桑人,科萨语虽然是以班图语为主干,似也混入了许多桑人典型的喀嗒声。在今日科技突飞猛进的时代,桑人在遗传与文化上的完整性不太可能再存续多久。幸好过去数十年来有不少研究,其目的都在于了解和记录这个独特的民族和他们的生活方式。约翰内斯堡威特沃特斯兰德大学(University of Witwatersrand)的托比亚(Philip Tobias)不仅是这些研究计划的发起人,多年来,在种族隔离的黑暗时期,他也担任桑人的非正式发言人,捍卫他们的权益。口悬河的威尔士人杰金斯(Trefor Jenkins)先是在赞比亚的数个铜矿城镇行医,后来搬到南非,他也是长期在遗传学方面研究桑人和其他原住民族群的先锋。
遗憾的是,即使最先进的遗传学技术,目前也仍无法理清人类文化的起源。考古证据显示,在进化的第一个阶段,我们的祖先的日常活动和其他的类人差不多,包拈尼安德特人在内。以色列的斯虎尔(Skhul)有一个洞穴遗址,其中的证据显示,大约在10万年前,现代智人和尼安德特人的族群曾经共同存在,没有任何一方对另外一方造成明显的威胁。但是,如同先前所见,现代人在3万年前左右消灭了眉脊突出的表亲尼安德特人。由此看来,似乎在这7万年期间,现代人因为技术以及(或者)文化上的进步而取得了优势。
个别的考古资料也支持这样的假设。显然大约在5万年前,现代人突然在“文化上”观代化了:我们从这个时期的遗物中发现,他们首度有了纯粹用于装饰的饰品,开始常规性地使用骨头、象牙和贝壳来制作日常的实用工艺品,并且也开始改良和采集的技术。当时究竟发生了什么事?我们可能永远无法知道。但我们忍不住猜想,或许是语言的发明促成了这一切,以及人类至今所有的成就。
根据定义,史前时代是指有书写纪录以前的时期,但是我们却发现,我们祖先的个别旅程早就记录在了我们每个人的DNA序列中。新兴的分子人类学运用不同族群的遗传变异模式来重建人类的拓殖史,借以窥知人类的“史前时代”。
研究遗传变异在各大洲的分布情况,再加上考古信息,提供了我们一些人类祖先在全球扩张的细节。人类沿着亚洲边缘,从如今的印度尼西亚群岛、新几内亚到澳洲的迁徙旅程,大约是在6万年前完成的。要抵达澳洲,必须横越数个浩大的水域,这显示我们的祖先在这个早期阶段已经开始使用船只。现代人类大约是在4万年前抵达了欧洲,又在大约1万年后越过北亚,包括日本。
亚利桑那大学的汉默(Michael Hammer)和这个领域的许多大师(包括肯恩和帕博)一样,也是在威尔逊的伯克莱实验室接受训练。虽然汉默最初感兴趣的是老鼠,但肯恩和威尔逊发表的线粒体DNA研究,让他从啮齿动物转向研究人类的历史。Y染色体的信息对肯恩和威尔逊的整个假说是很重要的试验,他是最早了解到这一点的人员之一。但是起初Y染色体不愿轻易让人窥知它的秘密。由吉尔伯特实验室所作的一项研究,把取自许多人Y染色体的同一个DNA区段定序,结果只发现这些区段的序列一模一样,辛苦研究半天,却没获得任何有关遗传上相互关系的信息。然而汉默坚持研究下去,最后他和其他人一起把Y染色体变成人类学的金矿,他们的成就在安德席尔发表那篇重要论文时达到了颠峰。
Y染色体的重要信息让我们得以重建人类拓殖新世界的历史,在整个人类史上,拓殖新世界是相对比较后期的发展阶段。目前有关人类最早在美洲的定居地点的说法仍莫衷一是,传统上获得最多支持的地点是新墨西哥州的克洛维斯(Clovis),时间约在1.12万年前;但是支持智利蒙特维尔第山(Monte Verde)的人则声称,那里的遗址至少已经有1.25万年之久。第一批美洲印第安人是在最后一次冰河时期穿过了白令海峡的陆桥,还是乘船走得比较偏南的路线,也仍有争议。然而,遗传数据明确显示,最早的创始族群很小:由于只找到两种主要的Y染色体序列,因此当时似乎只有两个明确的群体,而且每个群体可能顶多只有一个家庭。在印第安人之间,线粒体DNA变异比Y染色体变异广泛得多,显示每个创始群体中女性多于男性。在这两种Y染色体序列中,比较常见的那一种可能代表第一个创始群体;在第二个群体(包括今日纳瓦霍[Navajo]和阿帕切[Apache]的祖先)抵达前,第一个群体已建立了后代族群。比较常见的序列也具有另一个特色:它有一个在世上其他地方极少发现的突变。这个突变是2002年首度发现的,它进一步证明了其拥有者是更早的拓荒先驱,据估计,它已有1.5万年之久,略早于已知最古老的先民遗址。
遗传分析也让我们得以重建比较后期的史前历史。例如汉默已经证明现代的日本人,是古代以采集/渔猎为生的绳纹族(Jomon)和相对比较近代的弥生族(Yayoi)混种后的结果。绳纹族目前的代表族群是虾夷族(Ainu)原住民;弥生族大约在2500年前从朝鲜半岛抵达日本,带来了编织、金属工艺,以及以稻米为主的农业。我们也在欧洲看到一波波迁移的证据,这些迁移大多与农业技术的进步有关。巴斯克族(Basques)和凯尔特族(Celts)在遗传上与欧洲其余的族群不同。有一个解释是,这些族群面对更晚抵达的族群时,被迫迁移至相对较偏远的地区。(巴斯克人现在居住于法国和西班牙边境的比利牛斯山脉山区;凯尔特人较晚抵达欧洲,分布于欧洲西北缘,从法国的布列塔尼[Brittany]一直到爱尔兰和不列颠西部地区。)
牛津大学的塞克斯(Bryan Sykes)作了许多研究,揭露出现代欧洲复杂的遗传地图。传统看法向来认为,现代的欧洲人大多源自在地中海与波斯湾之间“肥沃的新月地带”(Fertile Crescent)、发明了农业的中东族群。但是塞克斯发现,欧洲人大多数的祖先不是源自于新月地带,而是源自于中东人入侵前欧洲本地更古老的族群,以及从欧亚中部(Central Eurasia)迁移过来的族群。这类族群包括分别在公元前500年及公元400年左右,从东方横扫欧洲的凯尔特人及匈奴人。塞克斯也在进一步阐述他对线粒体DNA的分析结果时,主张所有欧洲人都起源于7个“夏娃之女”(daughters of Eve)之一。所谓的“夏娃之女”是他对欧洲线粒体DNA族谱上少得惊人的几个主要祖先的称呼。他还成立了牛津寻祖公司(Oxford Ancestors),你可以付一笔钱,请该公司帮你定序你一部分的线粒体DNA,查清楚你究竟系出这“七仙女”中的哪一女。
了解人类史的另一个关键,可能在于卡瓦利-斯弗扎等人发挥得淋漓尽致的一项观察结果:遗传进化模式经常与语言进化模式有相互关系。当然,基因与语言之间明显具有一些可以模拟之处:它们都是代代相传,都会历经变化。但语言的变化可能特别快,青少年的父母应该都体会得到这一点。同样地,美式英语和英式英语相同,但是又具有明显不同之处,尽管它们才各自进化了几百年而已。因此,我们可以按照各个语言的异同点来重建语言族谱,这跟重建遗传族谱的方式大同小异。但是在许多实例中,如同达尔文本人所预测的 ,更重要的一点在于我们可以找出这两个族谱之间有用的对应关系,如此一来,我们对一个族谱的了解可以加深我们对另一个族谱的了解。凯尔特族和巴斯克族就是绝佳的实例:这两个族群在遗传上与欧洲的其余族群不同,其各自的语言也与欧陆其余的语言不同。至于在新世界方面,一个具有争议性的语言学理论指出,美洲只有三个本土的主要语族,其中两个与用印第安人的Y染色体资料辨识出来的早期迁徙事件有关;第三个语族是三者中最小的语族,则与孤立的因纽特人(Inuit,即爱斯基摩人)有关。
,更重要的一点在于我们可以找出这两个族谱之间有用的对应关系,如此一来,我们对一个族谱的了解可以加深我们对另一个族谱的了解。凯尔特族和巴斯克族就是绝佳的实例:这两个族群在遗传上与欧洲的其余族群不同,其各自的语言也与欧陆其余的语言不同。至于在新世界方面,一个具有争议性的语言学理论指出,美洲只有三个本土的主要语族,其中两个与用印第安人的Y染色体资料辨识出来的早期迁徙事件有关;第三个语族是三者中最小的语族,则与孤立的因纽特人(Inuit,即爱斯基摩人)有关。
在有了这些性别特异性(sex-specific)遗传资料(女性的线粒体DNA,男性的Y染色体)之后,就可以比较男性与女性的历史。卡瓦利-斯弗扎的研究生西斯塔德(Mark Seielstad)挑的题目是比较两性的迁移模式。这个研究的逻辑很简单:假设在南非的开普敦市,有一个Y染色体发生突变,则该突变传播至开罗的速度,就可以作为男性迁移速率的指标。同样地,在开普敦市出现的线粒体DNA突变传播至开罗的速度,可以用来测量女性迁移的速率。
不论好坏,历史向来是男性迁移的编年史,而非女性的。一般而言,男性出外是为了掠夺或建立帝国:亚历山大大帝从马其顿挥师入侵印度北部地区;维京人纵横大海,从斯堪的纳维亚到冰岛,甚至更远的北美洲;成吉思汗率领骑兵越过中亚的大草原等都是实例。但是即使没有战争作为旅行的原因,我们仍会认为在人类社会中,男性的移动性比较大。传统上男性负责狩猎,所以会到离家很远的地方,而在传统的釆集/狩猎社会,女性大多在家附近采集食物,养儿育女。西斯塔德因此认为,男性应该是传播人类基因的原动力。研究结果的数据却证明他错得离谱:一般而言,女性的移动性比男性大八倍!
事实上,这个模式虽然与我们的直觉印象相反,却很容易解释。几乎世界各地所有的传统社会都有人类学家所谓的父权地域制(patrilocality):来自不同村庄的人结婚后,女性会搬到男性所在的村庄,但男性不会搬到女性原本居住的村庄。假设现在有一个来自A村庄的妇女嫁给B村庄的人,然后她便搬到B村庄去,生了一个女儿和一个儿子。后来,这个女儿嫁到C村庄,这个儿子则把D村庄的妇女娶过来。由此看来,男系一直待在B村庄,但女系在两代之间从A移到B,再移到C。这个过程代代持续,结果女性的迁移范围很广,但男性的迁移范围则很小。男性的确偶尔会远离家乡去征服远方的土地,但是在人类浩大的迁移模式中,这些事件并不重要:至少在遗传的层面,人类历史其实是取决于女性在一个个村庄之间一步步的迁移。
针对一个区域,详细地研究该地Y染色体和线粒体DNA的变异,也可以看出在殖民过程中所造成的性关系与婚配习惯。比方说,在维京人抵达前,冰岛原本无人居住,但在对线粒体DNA与Y染色体进行比较后,我们发现有显著的不对称情形。大多数的Y染色体都如预期一样是来自斯堪的纳维亚人(Norse,通称北欧人),但大部分的线粒体DNA类型则是来自爱尔兰。斯堪的纳维亚人在殖民冰岛时,显然带着爱尔兰妇女。可惜我们无法从线粒体DNA的数据中得知当时爱尔兰妇女的感受。
最近一个针对哥伦比亚人Y染色体与线粒体DNA变异的研究,证明了类似的效应。在大多数的社会族群中,哥伦比亚人的Y染色体和西班牙人的Y染色体相同,这是征服南美洲北部沿岸西班牙属地的欧洲人留下来的生物遗产。事实上,在接受检测的Y染色体中,大约94%都是源自欧洲。似有趣的是,线粒体模式却相当不同:现代哥伦比亚人拥有多种类型的美洲原住民线粒体DNA。这样的结果所代表的含义相当清楚:入侵的西班牙人(当然都是男性)娶了当地的女人。哥伦比亚人没有美洲原住民的Y染色体类型,这透露出殖民时代的灭族大屠杀悲剧:当地的男性被消灭,而女性则被征服者通过性的方式“同化”了。然而,有时这种XY来源长久以来不对称的情况是因为文化延续的问题,而不是不同文化激烈冲突的结果。印度的少数民族帕西人(Parsee)认为自己是祆教徒(Zoroastrian)的后裔,祆教徒属于印欧民族雅利安人,在7世纪时因宗教迫害逃离波斯(今伊朗)。对现代帕西人所作的遗传分析显示,他们真的保有伊朗人的Y染色体,但他们的线粒体DNA通常是印度型的。在这个例子中,这种不对称性是因为传统而维持了下来:只有信奉祆教的帕西人所生的儿子,才会被视为真正信奉祆教的帕西人。因此帕西人社群里的成员都经由父系继承到Y染色体。遗传学确认了传统的影响力。
传统也与犹太人的遗传变异模式有关。最近一项研究已经证明,古代犹太人的祭司阶级kohanim及其后代(至今仍可由其姓氏柯恩[Cohen]来辨别)所拥有的Y染色体相当独特,足以使他们自成一个族群。即使居住地区最偏远的犹太族群,也就是在古代犹太人被逐出故土的“大流散”(Diaspora)中迁移至最远方的犹太族群,例如南非的兰巴(Lemba)。柯恩型Y染色体仍保存下来,几乎就像神圣的宗教经文一样。一般认为这种染色体的来源始于亚伦(Aaron);根据圣经记载,亚伦是祭司阶级的创始人,也是摩西的兄长。柯恩Y染色体序列真的源自亚伦,并在父子代代相传下保持完整,也并非不可能的事。在犹太人的历史中,这样的传统一直被严格保存下来。
到纽约市中心曼哈顿的街道上逛一圈,看到形形色色的人,你会觉得我们人类是地球上遗传变异最多的物种。但是,以我们已取得遗传信息的物种而言,相较于其中的大多数,其实人类基因组的变异明显少得多。就个体之间的差异而言,每1000个人类碱基对中大约只有1个是不同的。因此在遗传上,我们有99.9%相同,以其他物种的标准来看,这种差异算是很小的。尽管果蝇看起来都一样,但它们的变异程度却比我们高10倍。即使是南极洲上每一只都长得一模一样的阿德利企鹅(Adélie penguin),遗传变异程度也是我们的2倍以上。就连跟我们关系更近的黑猩猩,遗传变异程度也没我们这么少:黑猩猩的遗传变异程度大约是我们的3倍,大猩猩是2倍,红毛猩猩则是3.5倍。

亚伯拉罕的两个妻子各有一子,据说犹太人和阿拉伯人便是这对同父异母兄弟的后代。
汉默和其他人利用Y染色体来追踪犹太人的大流散过程,获得许多有趣的结果。例如在欧洲定居了1200年(现在也居住在美国和其他地方)的徳系犹太人(Ashkenazim),至今仍具有源自中东的遗传特征。事实上,分子研究已清楚显示,至少在遗传上,犹太人和所有其他的中东族群(包括巴勒斯坦人)毫无区别,经书上也是这样记载的。据说伟大的希伯来人族长亚伯拉罕(Abraham)跟不同的妻子生育了两名儿子:以撒(Isaac)和以实玛利(Ishmael)他们两人分别是犹太人和阿拉伯人的祖先。同一人的后代竞然成为不共戴天的死敌,这在基因证实了传统说法后,更显讽刺。
在获得线粒体DNA和Y染色体的家族数据后,我们人类这么相似的原因就昭然若揭了:我们共同的祖先存在的年代离现在并不远;就进化的标准而言,15万年的时间眨眼即过,在这么短的时间内,是没有办法经由突变产生大量丰富变异的。
关于人类的变异还有一个与我们的直觉极相反的发现:无论这些变异有多少,大多数都跟种族没有直接的关连。在肯恩和威尔逊证明人类离开非洲的时间出乎意料地晚之前,一般认为,不同的族群已在不同的大陆上互相隔离了无数年代,甚至长达200万年。依照泡令-祖克坎德尔模型,孤立的族群之间遗传变异的程度,跟他们隔绝的时间长短有连带关系,如此一来应该会累积大量的遗传差异。但是,根据肯恩和威尔逊的结论,我们共同的祖先生存的年代离现代并不远,所以地理位置相互隔离的族群显然没有足够时间发展出大量变异。因此,尽管肤色等遗传差异在不同族群之间显得很明显,但跟特定种族有关的遗传差异一般非常有限(即界定族群之变异并不多)。事实上,我们少量的变异大多相当均匀地分布在不同族群之间:我们在某个非洲族群找到特定遗传变异的几率,跟在某个欧洲族群找到相同变异的几率差不多。这让人不禁猜测,人类大多数的遗传变异是在离开非洲之前,就已经在非洲大陆发生,因此拓殖到世界其他地方的族群,才会都有这些变异。
就算我们对人类的遗传多样性还有一丁点自豪,也会在得知下面的事实后消散:人类基因组计划的结果显示,我们的DNA中大约只有2%为基因编码,这意味我们的变异至少有98%是发生在没有作用的基因组区域。由于自然选择非常有效率地消灭了对基因组中重要功能部分(例如基因)造成影响的突变,因此突变通常会在非编码的“垃圾”区域累积。我们之间的遗传差异很少,而这所造成的不同甚至更少。
由于人类的进化可说历时甚短,我们在族群中所看到的具有一致性的差异,大多数都可能是自然选择的产物,例如肤色。
黑猩猩是亲缘关系跟我们最近的动物,它们浓密毛发下的皮肤大多都没有色素——黑猩猩可以说是拥有一身白皮肤。在500万年前,人类和黑猩猩的共同祖先可能也同样是白皮肤,因此我们可以推论,现在非洲人(以及出生在非洲的最早的智人)特有的深肤色是在后来人类的进化过程中出现的。在失去毛发后,色素成为保护皮肤细胞不受阳光中紫外线辐射伤害的要素。
根据我们对分子的了解,现在我们已经知道紫外线会引起皮肤癌:它们会让双螺旋的相邻胸腺嘧啶碱基黏在一起,使DNA分子就像打结一样。当DNA进行复制时,这个结经常会造成碱基插错,从而造成突变。如果在偶然的情况下,那个突变刚好发生在调节细胞生长模式的基因上,就可能造成癌症。皮肤细胞产生的黑色素(melanin)能减少紫外线的伤害。跟我一样白皮肤的人都很清楚,虽然晒伤通常并不会致命,其对健康的威胁却比癌症大得多。因此也不难想像自然选择会有利于肤色变黑,这样不仅能防止癌症,也能预防严重晒伤可能引起的感染。
住在纬度较高的地区,为什么会使人失去黑色素?最好的解释是,这与维生素D3的合成有关,D3的合成过程是在皮肤内进行,而且需要紫外线。D3是钙质摄取的重要成分,而钙质则是使骨骼强的重要成分。(D3不足会造佝偻病和骨质疏松症。)在我们的祖先离开非洲到季节分明的环境后,全年的紫外线辐射比较少,所以自然选择有可能对肤色较白的变异有利,因为他们的皮肤内阻挡阳光的色素较少,在可用的紫外线有限的情况下,能以较高的效率合成D3。相同的逻辑也适用于我们在非洲大陆上迁移的祖先,例如南非的桑人,南非的紫外线强度跟地中海差不多,桑人的皮肤却特别白。似是住在几乎没有阳光的北极和附近地区、肤色却深得令人惊讶的因纽特人,又要怎么解释呢?由于气候因素,他们必须随时用衣服包住全身,这应该会进一步缩减他们制造维生素的机会。但是,事实上,在他们身上似乎看不到对浅肤色有利的自然选择压力,原因可能在于他们已经自行解决D3的问题:他们的饮食中富含鱼肉,而鱼肉正是这种重要养分的丰富来源。
虽然肤色在人类历史和个人生命上向来是很强大的决定因素(而且大多是负面的),但我们对肤色背后的遗传学,所知却少得令人惊讶。然而这可能不是因为科学本身的限制,而多半是政治介入科学的结果。在受到政治正确性压制的学术界,即使研究肤色这类特征的分子基础都是禁忌。目前我们在这方面的少许知识,都是来自以前对混血儿的研究,在这些研究中发现了数个促成色素沉着的基因。但是我们对其他物种的了解,以及所有哺乳动物之间基本生化过程的相似性都显示,真正的全貌没这么简单。例如,我们已经知道许多基因会影响老鼠的毛皮颜色,而且人类可能也有对等的基因。不过到目前为止,我们只找出了两个与人类色素沉着有关的基因:一个在发生突变时会引起白化症,另外一个是与红发和肤色淡(经常有雀斑)有关的黑皮质素受体(melanocortin receptor)。黑皮质素受体基因在欧洲人和亚洲人当中都有变异,但在非洲人中没有变异,这意味着,在非洲,自然选择对这个基因的突变非常不利,亦即不会选择让红发、皮肤白皙的人产生。完全缺乏色素的白化症患者,如今偶尔会在非洲族群中出现(可能是通过新生突变[de novo mutation]),但是由于他们对阳光极度敏感,所以处于非常不利的地位。
另一个可能也是由自然选择决定的形态性状是体型。在以散发体热为优先考虑的热带气候区,进化出了两种基本体型。一是高而痩的尼罗河流域体型(Nilotic form),以东非的马萨伊人(Masai)为代表,这种体型是把表面积对体积的比例增加到最大,以便促进散热。另一是侏儒体型(Pygmy form),也是细痩型,但很矮,矮小的体型比较适合需要耗费体力的采集/狩猎生活,使行动时耗费的能量减到最少。毕竟,何必拖着庞大的身躯去寻找食物?相对地,在高纬度地区,自然选择对能留存体热的体型有利,亦即表面积对体积的比例较低的体型。因此欧洲北部的尼安德特人身材粗壮,如今北方国家的居民一般也是这样。今日不同族群在运动表现上的差异,大概也可以归因于这些体型上的差异,例如在跳高方面,高大的尼罗河流域体型比矮壮的体型更适合,应该也没什么好奇怪的了。

体型对气候的进化适应。肯尼亚已经适应炎热气候的马萨伊人(左),以及格陵兰已经适应寒冷气候的因纽特人(即爱斯基摩人)。
若说有哪种性状在人类族群中的分布情况是很难了解的,那就是乳糖不耐症(lactose intolerance)。哺乳动物的乳汁,包括人类的,都富含一种糖类,称为乳糖;而初生的哺乳动物一般会制造一种特殊的酶,即乳糖酶(lactase),它会分解肠道中的乳糖。但是在断奶时,大多数哺乳动物,包括人类(至少包括大多数非洲人、美洲原住民和亚洲人),会停止制造乳糖酶,因此成人无法消化乳糖。有乳糖不耐症的人喝一杯牛奶就会引起腹泻、肠气和腹胀等不适症状。但相对地,大多数高加索人和其他一些族群的人却终生都会制造乳糖酶,因此终生都可以食用乳制品。
有一种解释指出,向来主要仰赖乳制品的族群进化出了乳糖耐受性(lactose tolerance)。但这个性状的模式仍无法令人完全信服这种解释,比方说,在中亚以畜牧为生、人人都以奶酪为食的族群,仍有乳糖不耐症。尽管我本身所属的族群通常可以耐受乳糖,但我本身却有乳糖不耐症。如果自然选择对特定族群的乳糖耐受性有利,为什么同一族群里不是所有的人都可以耐受乳糖?到目前为止,支持这种标准解释最有力的证据在于传统上以畜牧为生的非洲族群中,确实具有乳糖耐受性。我们可能永远无法完全了解这个性状的适应范围,但是研究芬兰族群的分子生物学家最近已经找出造成这个性状的突变。虽然这不是攸关生死的问题,但是现在只要做个简单的遗传试验,就可以得知一个新生儿在长大后有什么选择:终生不吃冰淇淋,还是长期忍受胃痛?
若说人种之间相对较少的差异很有趣,那么人类整体的共同特色就更有趣了:我们和亲缘关系最近的生物为什么会有这么大的差异呢?我们先前已经看到,人类大约在500万年前和黑猩猩分道扬镳,但是这段时间仅仅只够让我们进化出1%的遗传差异。不过就在这1%的差异中,有一些关键性的突变让我们成为如今这种会思考、会说话,了不起的生物。其他物种是否具有某个程度的思想能力,尚待讨论,但是其他物种都没有产生达芬奇和克里克这类人物,却是不争的事实。
人类和黑猩猩的染色体非常相似。但是黑猩猩有24对染色体,我们则是23对。近来我们发现,人类的第2号染色体是由黑猩猩的两个染色体结合而成。人类和黑猩猩的第9号染色体(人类的比较大)以及第12号染色体(黑猩猩的比较大)也有一些差异,染色体中的数个倒位(inversion,基因的排列顺序反转)也有所不同。这些染色体上的差异是否具有重要意义,目前还很难讲。
在生化层面,我们也不是很清楚人类有哪些相对的优点,目前为止,我们只知道人类和黑猩猩之间有两个差异。第一个差异是:两个物种的细胞外围都具备的一种糖分子,即唾液酸(sialic acid),在黑猩猩体内,这个分子通过一种酶的作用,会有些微变化;但是在人类体内,为这个酶编码的基因总是发生突变,亦即没有酶生成,而人类细胞表面的唾液酸也就没有变化。这个差异是否具有任何重要意义,我们不得而知。第二个差异在于FOXP2基因。这个在2002年由帕博的小组所发现的差异,可能更有意义。目前已知FOXP2基因与人类的语言有关。(由于已经发现FOXP2基因发生突变会引起语言障碍,因此这个基因被媒体称为“文法基因”[the grammar gene],但这个词容易引起误解。)在FOXP2蛋白质有715个氨基酸的链上,人类只有两个氨基酸和黑猩猩及大猩猩不同,黑猩猩和大猩猩的FOXP2蛋白质则是完全相同的。事实上,这些氨基酸在所有试验过的哺乳动物中都是一样的,只有人类不同。此外,对这个基因内部及周围的DNA变异模式所进行的统计分析显示,在人类进化期间,自然选择可能对此蛋白质进行了某种塑造。这让人不禁产生一个(尚且言之过早的)想法:FOXP2可能是进化上的确凿证据,就像枪击案中还在冒烟的枪,让人得以窥知语言起源上的关键步骤。
帕博的实验室也率先釆取一项看来颇有希望的创新方法,寻找其他可能为重要差异编码的基因。帕博利用可决定特定组织内有哪些基因被激活的DNA微阵列(参见第八章),比较人类、黑猩猩和猕猴体内三种组织——白血球、肝组织和脑组织——的基因表现模式(亦即哪些基因被启动)。人类和黑猩猩的亲缘关系近,结果发现人类与黑猩猩的白血球和肝组织在基因表现上也很接近,如同预期。但脑组织的基因表现却截然不同:人类的脑和黑猩猩及猕猴的脑非常不同。或许这也没什么好惊讶的,大多数的人不需要有大堆仪器的实验室,也知道人脑和黑猩猩的脑显然不同。这个研究的意义在于提供在人脑与黑猩猩脑中表现不同的基因清单。但这也只是开端而已。就算找到基本机制的完整清单,我们也无法明确了解它们是“如何”使我们和黑猩猩变得不同。我们之所以为人,绝不是详细列出受控的分子就能说明的。是在搜寻人之所以为人的遗传基础之际,至少现在我们已经可以开始编纂一份包含各种可能原因的清单。
在我撰写本书时,黑猩猩基因组计划已经接近完成,等这个计划完成后,将会揭开金和威尔逊所找到的、构成那1%差异的DNA之秘密。我的猜想是,最终会证明他们的想法是对的:造成关键性差异的,不在于基因本身,而在于基因的调控。我怀疑,人类只不过是拥有几个独特(而且具有特定功能)的基因开关的大猿罢了。
分子生物学最伟大的任务当然是回答有关人类,以及人类这个物种起源的问题。但是,除了人类全体的故事,每一个人都会渴望知道其本身的故事。DNA也可以提供更个人化的谱系记录,让我们更加了解我们各自的祖先。我的DNA分子内可以说是记录着我的进化传承历史,那是一个可以从不同层面来看的故事。我可以把我的线粒体DNA序列放进肯恩和威尔逊的人类族谱,或者我也可以更仔细地了解自己家族的过去。我的Y染色体和线粒体DNA将述说不同的故事:一个说的是我父亲那边,一个说的是我母亲那边的故事。
我以前对族谱学丝毫不感兴趣。但我的家族(我猜许多家族都一样)有一位专门的档案保管人,那就是我的贝蒂阿姨。她把一生的时间都花在了解家族成员之间的关系,以及这些关系的来龙去脉上。她发现我们沃森家(源自苏格兰低地家族)是1795年首度抵达美国,在新泽西州的卡姆登市(Camden)落户。她也坚信,林肯在伊利诺伊州春田市的宅第是我父系某位祖先所设计的。但我向来对我的爱尔兰血统比较感兴趣,也就是我外婆的家族。我的外曾祖父母在19世纪40年代马铃薯大饥荒期间逃离爱尔兰,最后来到印第安纳州。我的外曾祖父葛利森(Michael Gleason)于1899年过世,同年我母亲诞生。在我外曾祖父的墓石上刻着,他来自爱兰尔的格雷(Glay)镇。
有一次到爱尔兰时,我特别到蒂珀雷里郡(Tipperary)的档案室,寻找更多与我外曾祖父有关的资料。那个档案室位于尼纳(Neneagh),离利默里克郡(Limerick)大约20英里,先前是一所监狱。但我的侦探工作一无所获,那里完全找不到有关格雷镇的记录,我只能判断说,在我那位可能不识字的祖先的墓碑上所刻的市名是想像出来的。因此,我结束了惟一一次针对自己家谱的研究。一直到最近,我对族谱的兴趣才又被勾起来。现在人类族谱的架构已经由肯恩和其他人建立起来,我很想知道自己在人类族谱里的位置。塞克斯的“牛津寻祖”这类公司,代表了家谱研究的新领域。他们用高科技的实验室取代布满灰尘的档案。牛津寻祖公司在取得我的DNA样本后,开始进行线粒体DNA和Y染色体的分析。只可惜检验结果显示,有关我祖先的历史一点也不浪漫,他们也不是来自遥远的异国。我真的跟我所害怕的一样,主要是苏格兰人与爱尔兰人的产物。我甚至不能把我比较野蛮的一面,归咎于古代时曾有维京人侵入我的家族血统。
桑人也称为布须曼人(Bushmen,荷兰语为Sanqua,意为“丛林人”。这是17世纪晚期定居南非的荷兰移民对他们的轻蔑称呼。
达尔文在《物种起源》中提到:“如果我们有完整的人类族谱,人种的族谱排列可以提供现今世上多种语音的最佳分类方法。”